
Nymphalidae de la Caldera de Lubá. Isla de Bioko (Guinea Ecuatorial) (Lepidoptera: Nymphalidae)
Nymphalidae of the Caldera de Lubá. Bioko Island (Equatorial Guinea) (Lepidoptera: Nymphalidae)
Nymphalidae de la Caldera de Lubá. Isla de Bioko (Guinea Ecuatorial) (Lepidoptera: Nymphalidae)
SHILAP Revista de lepidopterología, vol. 44, núm. 175, 2016
Sociedad Hispano-Luso-Americana de Lepidopterología
Recepción: 10 Octubre 2014
Aprobación: 24 Noviembre 2014
Resumen: En el presente trabajo se presentan los primeros registros de los Nymphalidae de la Caldera de Lubá (Isla de Bioko). Siete de las 46 especies mostradas en el presente trabajo, Cymothoe althea, C. consanguis, C. cape- lla, Lachnoptera anticlia, Amauris vashti, Bicyclus golo . Bicyclus neustetteri representan las primeras citas pa ra Bioko. De estas especies sólo Lachnoptera anticlia, Amauris vashti . Cymothoe capella aparecen citadas en la región continental, pero para el resto también suponen las primeras citas para Guinea Equatorial. Se usaron diversas medidas de riqueza y diversidad (Margalef, Simpson, Shannon-Weaver), mostrando todos ellos altos valores.
Palabras clave: Lepidoptera, Nymphalidae, Caldera de Lubá, riqueza de especies, estimas de diversidad, curvas de acumulación, Bioko, Guinea Ecuatorial.
Abstract: The present paper shows the first records of butterflies Nymphalidae of the Caldera de Lubá (Bioko Island). Seven from the 46 species showed, Cymothoe althea, C. consanguis, C. capella, Lachnoptera anticlia, Amauris vashti, Bicyclus golo and Bicyclus neustetteri, represent the first cites of Bioko. From these species only Lachnoptera anticlia, Amauris vashti and Cymothoe capella are cited in Continental Region, but for others also represent the first cites of Equatorial Guinea. Several measures of richness and diversity were used (Marga- lef, Simpson, Shannon-Weaver), showing all high values.
Keywords: Lepidoptera, Nymphalidae, Caldera de Lubá, species richness, diversity estimations, accumula- tion curves, Bioko, Equatorial Guinea.
Introducción
El origen, la diversificación y la taxonomía de la familia Nymphalidae está siendo revisada en los últimos años, tanto desde el punto de vista evolutivo (ZHANG et al., 2008) como filogenético (FREITAS & BROWN, 2004) y sistemático (WAHLBERG et al., 2003). Está consensuado que la familia Nymphalidae mantiene un origen común con las otras cuatro familias de Papilionoidea, to- das ellas originadas y diversificadas en el Terciario (VANE-WRIGHT, 2004). Actualmente se incluye en esta familia una importante diversidad de subfamilias, muchas de ellas consideradas hasta fe- chas recientes como familias independientes -Satyridae, Acreidae, etc.-, pero diversos estudios evolutivos y filogenéticos del grupo han evidenciado su pertenencia a los ninfálidos (BROWER, 2000; ZHANG et al., 2008; WAHLBERG et al., 2009). La clasificación de las subfamilias elaborada por ACKERY et al., (1999), seguida desde entonces por muchos autores, se ha mantenido vigente hasta fechas recientes y han sido pocos los cambios realizados (BROWER, 2000), generalmente solo implicando a determinados géneros (Calinaga, Phyciodes, etc.) (WAHLBERG et al., 2003). En la presente década, los más recientes estudios sobre la familia han determinado algunas variaciones significativas en su organización en subfamilias (WAHLBERG et al., 2005), siendo comúnmente aceptada la clasificación presentada por MADDISON & SCHULZ (2007) y seguida en los trabajos generalistas actuales (WILLIAMS, 2008; SÁFIÁN et al., 2009), con algunas pequeñas modificaciones posteriores en los niveles inferiores (NYLIN et al., 2014). Así pues, esta será la clasificación seguida en este trabajo.
Aproximadamente la tercera parte de los ropalóceros del mundo pertenecen a los Nymphalidae (ZHANG et al., 2008), es decir, más de 6.000 especies de mariposas están incluidas en esta familia de las que alrededor de 1.500 viven en África subsahariana, ocupando por tanto la región afrotropical (ACKERY et al., 1995; WILLIAMS, 2008). El Oeste del continente cuenta con cerca de 600 especies de ninfálidos (LARSEN, 2005; TURLIN, 2007) de las cuales aproximadamente 140 están ci- tadas en Bioko (Martín et al., en prep).
La Isla de Bioko es considerada como una de las zonas geográficas de mayor biodiversidad a nivel mundial (BURGESS et al., 2006) lo cual, sumado a que alrededor del 90% de las mariposas diurnas conocidas viven en los sistemas tropicales (LARSEN, 1995; BONEBRAKE et al., 2010), hace de esta isla del Golfo de Biafra un lugar de extraordinaria diversidad de lepidópteros ropalóce- ros (SPEARMAN et al., 2000), particularmente de la familia Nymphalidae (HENNING, 1988; TURLIN, 1999; LARSEN, 2005; TURLIN, 2007), aun cuando la región sur, considerada como el área de mayor riqueza de especies de Bioko (BUTYNSKI & KOSTER, 1994; OBAMA, 2006), per- manece prácticamente inexplorada (NAVARRO et al., 2012). Así pues, entendiendo la diversidad de una comunidad como una expresión del reparto de recursos y energía, su estudio es una de las aproximaciones más útiles en el análisis comparado y una herramienta básica para su conservación (HALFFTER & EZCURRA, 1992). Por ello, tal como apunta SAMWAYS (1994) existe una auténtica necesidad de identificar áreas de máxima diversidad de insectos (‘Hot Spots’) y centros de en- demicidad o rareza.
Diversidad y riqueza son conceptos tradicionalmente unidos y, en la actualidad, es sumamente común asimilar el concepto diversidad -en sentido estricto- a una función de relación entre la riqueza y la abundancia relativa de sus elementos, en nuestro caso especies de ropalóceros de la familia Nymphalidae (HEYWOOD, 1994). Para medirla existen diferentes funciones e índices y aunque hoy día hay más de 60 índices que ayudan a cuantificar la diversidad, los más frecuentemente usa- dos son dos o tres: Shannon-Weaver, Simpson, Margalef, etc. Todos miden similares unidades, tra- tando de encontrar el ajuste entre riqueza y abundancia relativa de los elementos individuales (MORENO, 2001). No obstante y en caso de querer comparar localidades, su uso es mucho menos frecuente y el número de especies (Riqueza) ha pasado a ser el parámetro comúnmente utilizado (GOTELLI & COLWELL, 2001). Así pues, partiremos del uso de “Especie” como medida de diver- sidad debido a tres factores primordiales: la riqueza (.) ya refleja distintos aspectos de biodiversidad, es decir, es un estimador sencillo de diversidad, denominada “diversidad alfa” (WHITTAKER, 1972); el significado de especie está ya consensuado y, al menos para los lepidópteros, en general las especies son detectables, cuantificables y hay suficiente información sobre su número (MORENO et al., 2011). De este modo, si entendemos la diversidad alfa como el resultado del proceso evolutivo que se manifiesta en la existencia de diferentes especies dentro de un hábitat particular, entonces un simple conteo de .sería suficiente para describir esa diversidad sin necesidad de evaluar el peso o valor de cada especie dentro de la comunidad (MARGALEF, 1958), pero lo más conveniente es presentar valores tanto de la riqueza como de algún índice de la estructura de la comunidad, de tal forma que ambos parámetros sean complementarios en la descripción de diversidad (NÚÑEZ, 1991; MORENO, 2001; MORENO et al., 2011).
Por otra parte, es evidente que resulta imposible registrar la totalidad de especies en un inventario, más aún al tratarse de mariposas en sistemas tropicales (GOTELLI & COLWELL, 2001) al ser éstos los lugares de mayor diversidad de ropalóceros del planeta (BONEBRAKE et al., 2010). Para este tipo de trabajos las funciones de acumulación de especies resultan efectivas para predecir las especies esperadas que se obtendrían con un tamaño mayor de muestreo (VANE-WRIGHT et al., 1991). Son usadas, por tanto, para dar fiabilidad a los inventarios biológicos y posibilitar su comparación. Estas funciones son habitualmente aplicadas en estudios con diversos grupos, tanto invertebrados (JIMÉNEZ-VALVERDE & HORTAL, 2003; ZAMORA et al., 2011) como vertebrados (GONZÁLEZ-OREJA, 2010; MARTÍN et al., 2014). Este análisis permite identificar la diversidad biológica máxima en un determinado territorio, lo que es un requisito primordial para la conservación (VANE-WRIGHT et al., 1991; SCOTT, 1997).
En el presente trabajo se muestran los ropalóceros de la familia Nymhalidae recolectados en el interior de la Caldera de Lubá durante las expediciones realizadas en 2005 y 2007 por la Universidad Politécnica de Madrid, así como los resultados de la aplicación de índices de diversidad de distinto orden y la elaboración de curvas de acumulación de especies.
Materiales y Métodos
El área de estudio se encuadra en la Reserva Científica de la Gran Caldera y Tierras Altas del Sur de Bioko (Figura 1), representando la única muestra de bosque monzónico de Guinea Ecuatorial (NAVARRO et al., 2012). En la Caldera Volcánica de Lubá se abre un espectacular cráter de 5 km de diámetro, con desniveles de más de 1.400 m originados por el hundimiento de la antigua cumbre (FUSTER, 1956), constituyendo por ello un terreno extraordinariamente complejo, encerrado por paredes verticales repletas de selva y surcado por profundos y angostos barrancos (MARTÍNEZ, 1968; MARTÍN & COBOS, 2010). Estas condiciones justifican el estado prístino de su hábitat, inalterado y sin influencia humana en su evolución.
El trabajo de campo se realizó en diciembre de 2005 y marzo de 2007, contabilizándose un total de 26 jornadas de muestreo. Todas las capturas se obtuvieron, en hábitat de bosque monzónico primario, no perturbado. La altitud máxima de los registros fue de 1293 msnm y la mínima a nivel del mar. En la figura 2 se muestran las ubicaciones de las capturas, localizadas por sus correspondientes coordenadas y altitud.
Para la aplicación de índices de diversidad se usó el cálculo conceptual de diversidad alfa (WHITTAKER, 1972) ya que trabajamos en ambiente prístino -y por tanto no intervenido- de bosque monzónico continuo, pudiendo así hablar de Riqueza (S) como medida de diversidad (RICOTTA, 2005). De este modo, se aplican métodos basados en la cuantificación del número de especies (riqueza específica) y otros basados en la estructura de la comunidad, tanto fundamentados en la dominancia como en la equidad.
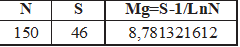
Para medir la Riqueza empleamos el Índice de diversidad de Margalef (DMg), expresado mediante la relación: DMg=.-1/ln., donde S es el número total de especies (riqueza específica) y N el número total de individuos. Este índice supone que hay una relación funcional entre el número de especies y el número total de individuos, de modo que el índice varía en función del tamaño de la muestra, considerándose la comunidad de escasa diversidad con valores inferiores a 2 y como muy diversa a partir del valor 5 (MARGALEF, 1958).
El índice de Simpson (l=åp.2) es en realidad un estimador de dominancia tipo “lambda” que, por definición, se opone a la diversidad. Por ello, a mayor valor de este índice menor será la diversidad. La expresión de la Diversidad es:
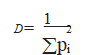
donde . representa la probabilidad de que dos individuos tomados al azar pertenezcan a la misma especie, P. representa la abundancia relativa de la especie i, es decir, n./N, siendo N= número total de ejemplares o población total. La mínima diversidad viene dada por un índice de valor 1, absoluta dominancia de una especie. De igual modo, índices de valores de “lambda” (l) muy pequeños implican comunidades muy diversas. Por último destacar que los valores de este índice son sensibles a las abundancias de una o varias de las especies más frecuentes de la comunidad y puede ser considerado como una medida de la concentración dominante (NÚÑEZ, 1991).
Posiblemente el índice de equidad de Shannon-Weaver es uno de los más usados en biología (BAEV & PENEV, 1995; MORENO et al., 2011) ya que expresa la uniformidad de los valores de importancia a través de todas las especies de la muestra. Se representa mediante la expresión H´=-åp.lnpi. En definitiva, este índice mide el grado de incertidumbre en predecir a que especie pertenecerá un individuo escogido al azar en una muestra (MAGURRAN, 1998) asumiendo, por tanto, que todas las especies están representadas en la muestra y los individuos son seleccionados al azar. Adquiere valores entre cero cuando solo hay una especie y el logaritmo de S cuando todas las especies presentan el mismo número de individuos (MORENO, 2001).
Como ya señalamos, el uso de curvas de acumulación de especies se muestra como una herramienta muy efectiva para estimar el número de especies esperado a partir de un muestreo. En el caso de invertebrados esos valores extrapolados, es decir la riqueza esperada, pueden ser usados como medida de la diversidad alfa, evitando así los sesgos que se establecen en los inevitables submues- treos cuando se trabaja con insectos (ESPINOSA, 2003; BECK & SCHWANGHART, 2010). Los modelos y expresiones matemáticas comúnmente utilizados son los modelos lineales o los métodos no paramétricos. Los primeros son usados cuando se asume un tipo de distribución estadística (matemática) conocida (JOST, 2010), mientras que los segundos se ajustan a funciones no-paramétricas en sentido estadístico, pues no asumen el tipo de distribución del conjunto de datos y no los ajustan a un modelo determinado, requiriendo únicamente datos de presencia/ausencia (MORENO, 2001).
MODELOS LINEALES
La ecuación de De Clench, adaptada de la ecuación original de Michaelis-Menten (MM Means), es un modelo comúnmente utilizado y ha demostrado un buen ajuste a los inventarios con ropalóceros (SOBERÓN & LLORENTE, 1993) y heteróceros (RICKETTS et al., 2004). No obstante, esta ecuación está recomendada para zonas de muestreo amplias y con protocolos que demandan cambios en la intensidad del esfuerzo de muestreo en campo (JIMÉNEZ-VALVERDE & HORTAL, 2003), de modo que se pueda asumir un patrón de ajuste de los datos (MORENO, 2001; MORENO et al., 2011) al contar con un gran tamaño de muestras y asumiendo, por tanto, que la riqueza total será el número de especies halladas tras un esfuerzo de muestreo infinito (MAGURRAN, 2007). Debemos destacar que este tipo de modelos han resultado eficaces para dimensionar el esfuerzo en los trabajos de muestreo (COLWELL et al., 2004; JOST, 2010). Su expresión matemática recomendada es: S.=.n/1+.n, donde . representa la tasa de crecimiento de nuevas especies y . es un parámetro relacionado con la forma de la curva, siendo n el número acumulativo de muestras. Aunque este tipo de modelos no son recomendados para las características de nuestros registros, usaremos su aplicación como medida del esfuerzo necesario para completar el inventario hasta el límite señalado por los estimadores que mejor se ajusten a nuestros datos (MORENO & HALFFTER, 2001).
MODELOS NO PARAMÉTRICOSMODELOS NO PARAMÉTRICOS
Cuando no disponemos de datos del número de individuos pues no conocemos cómo se comporta la distribución de los individuos por especie, las funciones que mejor se ajustan están basadas en modelos no-paramétricos (MORENO, 2001; BECK & SCHWANGHART, 2010; GONZÁLEZ- OREJA et al., 2010). Entre los modelos no paramétricos, los estimadores utilizados en el presente trabajo son Jacknife 1 y 2 (MORENO, 2001) y Chao2 (CHAO, 1984). Para todos ellos . representa el número de especies “únicas”, que ocurren solamente en una muestra; . es el número de especies que ocurren exactamente en dos muestras y . el número de muestras.
Jacknife 1 o de primer orden (cuya expresión es J.= S+L (m-1)/m) se basa en el número de especies de una muestra y reduce el sesgo de los valores estimados. Ha demostrado un buen ajuste como límite inferior en la estimación de especies potenciales (GONZÁLEZ-OREJA et al., 2010). Jacknife 2 o de segundo orden (J2= S+L (2m-3)/m - M(m-2)2/m(m-1)) se basa en el número de especies que ocurren en una muestra así como en el número de ellas que lo hacen en exactamente dos (PALMER, 1990). Dentro de este tipo de estimadores Chao 2 es el más riguroso y menos sesgado de todos (MORENO & HALFFTER, 2001; GONZÁLEZ-OREJA et al., 2010) y estima el número de especies esperadas, considerando la relación entre el número de especies únicas (que sólo aparecen en una muestra) y el número de especies duplicadas (que aparecen compartidas en, al menos, dos muestras) (CHAO, 2005) y su expresión matemática es Chao.=S+L2/2M. Este tipo de expresiones se pueden ajustar en cualquier programa estadístico (Statistica 12 o similar) con un procedimiento de regresión no lineal definida por el usuario (SOBERÓN & LLORENTE, 1993). Eliminamos el posible efecto del orden en que se añaden las muestras a la curva de acumulación mediante el remuestreo aleatorio de las unidades de muestreo, usando para ello 100 aleatorizaciones mediante el programa EstimateS 9.1.0 (COLWELL, 2006).
En todos los casos, para la elaboración de las funciones de acumulación de especies establecemos cada zona de muestreo -descritas en la figura 2- como unidades válidas para cuantificar el esfuerzo empleado en el inventario y construir, a partir de este dato, las correspondientes curvas de acumulación (MORENO & HALFTER, 2001).
Resultados
Del total de 70 especies registradas para el interior de la Caldera de Lubá, 46 son Nymphalidae, representando en conjunto alrededor del 65% del total. Las capturas se reparten en 9 subfamilias, siendo 9 Nymphalinae, 1 Charaxinae, 14 Limenitinae, 1 Cyrestinae, 2 Biblidinae, 8 Heliconii- nae, 4 Danainae y 6 Satyrinae. Todos los registros fueron colectados por la Expediciones Científicas-UPM a la Caldera de Lubá (Exp. UPM 2005 y Exp. UPM 2007).
NYMPHALIDAE NYMPHALINAE
Hypolimnas anthedon (Doubleday, 1845)
Campamento UPM, 1 1, XII-2005 (Exp. UPM 2005 leg.); Río San Antonio, 1 1, 13-III-2007 (Exp. UPM leg.); Río Riaco, 16-III-2007, 1 1 (Exp. UPM leg.).
Hypolimnas misippus (Linnaeus, 1764) Moraca, 1
1, 6-III-2007 (Exp. UPM leg.).
Hypolimnas salmacis insularis Schultze, 1920
Río Riaco, 1 1, 15-XII-2005 (Exp. UPM 2005 leg.); Río Riaco, 1 1, 9-III-2007 (Exp. UPM leg.); Campamento Ureka, 1 1, 12-III-2007 (Exp. UPM leg.); Pizarras, 1&&, 12-III-2007 (Exp. UPM leg.).
Protogoniomorpha parhassus (Drury, 1782)
Campamento Hormiga-Campamento UPM, 1 1, 8-III-2007 (Exp. UPM leg.).
Precis milonia Felder & Felder, 1867
Río Riaco, XII-2005 (Exp. UPM 2005 leg.); Poza Verónica, 16-III-2007 (Exp. UPM leg.); Campamento UPM-Campamento Hormiga, 18-III-2007 (Exp. UPM leg.).
Precis sinuata Plötz, 1880
Río Riaco, XII-2005 (Exp. UPM 2005 leg.); Campamento Ureka, 13-XII-2005 (Exp. UPM 2005 leg.); Río Riaco, 9-III-2007 (Exp. UPM leg.).
Kallimoides rumia (Doubleday, 1849)
Pizarras, 1 1, 12-III-2007 (Exp. UPM leg.); Río Riaco, 2 11, 16-III-2007 (Exp. UPM leg.).
Antanartia delius guineensis Howarth, 1966
Río Riaco, XII-2005 (Exp. UPM 2005 leg.); Río San Antonio, 13-III-2007 (Exp. UPM leg.); Río Riaco, 16-III-2007 (Exp. UPM leg.).
Antanartia dimorphica mortoni Howarth, 1966 Moraca,
1 1, XII-2005 (Exp. UPM 2005 leg.).
CHARAXINAE
Charaxes fulvescens marialuisae Canu, 1989
Campamento UPM-Campamento Hormiga, 1 1, 18-III-2007 (Exp. UPM leg.).
LIMENITINAE
Cymothoe althea (Cramer, 1776)
Campamento Ureka-Fondo Caldera, 1 0, 17-III-2007 (Exp. UPM leg.).
Cymothoe beckeri (Herrich-Schaeffer, 1858)
Río San Antonio, 1 0, 13-III-2007 (Exp. UPM leg.); Fondo Caldera, 1 0, 15-III-2007 (Exp. UPM leg.)
Cymothoe capella (Ward, 1871)
Río San Antonio, 3 1, 13-III-2007 (Exp. UPM leg.).
Cymothoe consanguis Aurivillius, 1896
Chapa Herminio, 1 1 y 1 0, 7-III-2007 (Exp. UPM leg.).
Cymothoe oemilius fernandina Hall, 1929
Pizarras, 1 1 y 1 0, 12-III-2007 (Exp. UPM leg.); Fondo Caldera, 3 1, 15-III-2007 (Exp. UPM leg.).
Cymothoe owassae Schultze, 1916
Fondo Caldera, 1 1 y 1 0, 15-III-2007 (Exp. UPM leg.).
Cymothoe caenis (Drury, 1773)
Pizarras, 1 1, 12-III-2007 (Exp. UPM leg.).
Catuna critea canui (Drury, 1773)
Campamento UPM, 8 11, 14-XII-2005 (Exp. UPM 2005 leg.); Campamento Ureka-Fondo Caldera, 1 0, 14-III-2007 (Exp. UPM leg.); Poza Verónica, 1 0, 16-III-2007 (Exp. UPM leg.).
Euphaedra canui Hecq, 1987
Río San Antonio, 1 0, 11-III-2007 (Exp. UPM leg.).
Pseudacraea eurytus (Linnaeus, 1758)
Pizarras, 1 1 y 1 0, 12-III-2007 (Exp. UPM leg.); Campamento UPM-Campamento Hormiga, 1 1, 18-III-2007 (Exp. UPM leg.).
Pseudacraea lucretia (Cramer, [1775])
Campamento Ureka, 1 0, 12-III-2007 (Exp. UPM leg.); Fondo Caldera, 1 1, 15-III-2007 (Exp. UPM leg.); Río Riaco, 3 00, 16-III-2007 (Exp. UPM leg.).
Pseudacraea semire (Cramer, 1779)
Campamento Hormiga-Campamento UPM, 8-III-2007 (Exp. UPM leg.); Fondo Caldera, 15- III-2007, 3 ex. (Exp. UPM leg.); Campamento UPM-Campamento Hormiga, 18-III-2007, (Exp. UPM leg.).
Euriphene incerta biokensis Hecq, 1994
Chapa Herminio, 1 1, 7-III-2007 (Exp. UPM leg.); Campamento UPM, 1 0, 8-III-2007 (Exp. UPM leg.); Río San Antonio, 2 00, 11-III-2007 (Exp. UPM leg.); Pizarras, 2 11 y 1 0, 12-III-2007, (Exp. UPM leg.).
Euriphene canui Hecq, 1987
Campamento UPM, 1 1 y 1 0, 8-III-2007 (Exp. UPM leg.).
CYRESTINAE
Cyrestis camillus (Fabricius, 1781)
Río Riaco, 9-III-2007 (Exp. UPM leg.).
BIBLIDINAE
Eurytela hiarbas (Drury, 1782)
Río San Antonio, 1 1 y 1 0, 11-III-2007 (Exp. UPM leg.); Río San Antonio, 1 0, 13-III-2007 (Exp. UPM leg.).
Mesoxantha ethosea ethoseoides Rebel, 1914
Río Riaco, 1 1, 15-XII-2005 (Exp. UPM 2005 leg.).
HELICONIINAE
Lachnoptera anticlia (Hübner, 1819)
Río Riaco, 1 1, 16-III-2007 (Exp. UPM leg.).
Acraea lycoa Godart, 1819
Chapa Herminio, 1 1, 7-III-2007 (Exp. UPM leg.); Pizarras, 1 0, 12-III-2007 (Exp. UPM leg.); Campamento UPM-Campamento Hormiga, 1 1 y 1 0, 18-III-2007 (Exp. UPM leg.); Chapa Herminio, 1 1, 19-III-2007 (Exp. UPM leg.).
Acraea umbra macarioides (Aurivillius, 1893) Río
Riaco, 1 0, 8-III-2007 (Exp. UPM leg.).
Acraea peneleos Ward, 1871
Campamento UPM-Río Riaco, 1 0, 8-III-2007 (Exp. UPM leg.).
Acraea circeis (Drury, 1782)
Campamento UPM-Campamento Hormiga, 1 0, 18-III-2007 (Exp. UPM leg.).
Acraea abdera Hewitson, 1852
Rio Riaco, 1 1, 8-III-2007 (Exp. UPM leg.).
Acraea pharsalus Ward, 1871
Campamento UPM-Río Riaco, 1 0, 8-III-2007 (Exp. UPM leg.).
Acraea epaea insulana Ackery, 1995
Campamento UPM, 1 1, XII-2005 (Exp. UPM 2005 leg.); Chapa Herminio, 1 0, 7-III-2007 (Exp. UPM leg.); Campamento Ureka, 1 1, 12-III-2007 (Exp. UPM leg.); Fondo Caldera, 3 11 y 1 0, 15-III-2007 (Exp. UPM leg.); Campamento UPM-Campamento Hormiga, 1 0, 18-III-2007, (Exp. UPM leg.).
Acraea excisa (Butler, 1874)
Río Riaco, 1 0, 16-III-2007 (Exp. UPM leg.).
DANAINAE
Amauris echeria fernandina Schultze, 1914
Río Riaco, 16-III-2007 (Exp. UPM leg.); Campamento UPM-Campamento Hormiga, 18-III- 2007 (Exp. UPM leg.).
Amauris inferna moka Talbot, 1940
Moraca, 3 ex., 6-III-2007 (Exp. UPM leg.); Campamento UPM-Campamento Hormiga, 8-III- 2007 (Exp. UPM leg.); Río Riaco, 4 ex., 9-III-2007, (Exp. UPM leg.); Fondo Caldera, 2 ex., 15-III- 2007 (Exp. UPM leg.); Río Riaco, 16-III-2007 (Exp. UPM leg.); Campamento UPM-Campamento Hormiga, 3 ex., 18-III-2007 (Exp. UPM leg.).
Amauris niavius (Linnaeus, 1758)
Campamento UPM, 1 1, XII-2005 (Exp. UPM2005 leg.); Fondo Caldera, 1 1, 15-III-2007 (Exp. UPM leg.); Campamento UPM-Campamento Hormiga, 2 11, 18-III-2007 (Exp. UPM leg.).
Amauris vashti (Butler, 1869)
Campamento UPM-Campamento Hormiga, 1 0, 18-III-2007 (Exp. UPM leg.).
SATYRINAE
Bicyclus golo (Aurivillius, 1893)
Campamento UPM, 2 11, 8-III-2007 (Exp. UPM leg.); Río San Antonio, 1 1, 11-III-2007 (Exp. UPM leg.); Campamento Ureka, 1 1, 11-III-2007 (Exp. UPM leg.); Fondo Caldera, 2 11, 14- III-2007 (Exp. UPM leg.); Fondo Caldera, 1 1, 15-III-2007 (Exp. UPM leg.)
Bicyclus neustetteri (Rebel, 1914)
Campamento UPM, 1 0, 8-III-2007 (Exp. UPM leg.); Campamento UPM-Río Riaco, 1 1 y 2 00, 8-III-2007 (Exp. UPM leg.); Pizarras, 1 1 y 1 0, 12-III-2007 (Exp. UPM leg.); Río San Anto- nio, 1 1, 13-III-2007 (Exp. UPM leg.); Campamento Ureka-Fondo Caldera, 1 0, 14-III-2007 (Exp. UPM leg.); Río Riaco, 1 1 y 3 (00, 16-III-2007 (Exp. UPM leg.); Poza Verónica, 1 0, 16-III-2007 (Exp. UPM leg.); Campamento UPM-Campamento Hormiga, 1 1, 18-III-2007 (Exp. UPM leg.)
Bicyclus dorothea concolor Condamin & Fox, 1964 Moraca, 2 11 y 1 0, 6-III-2007 (Exp. UPM leg.).
Bicyclus ignobilis eurini Condamin & Fox, 1963
Río Riaco, 2 00, 15-XII-2005 (Exp. UPM2005 leg.); Campamento Ureka-Fondo Caldera, 1 1, 14-III-2007 (Exp. UPM leg.); Poza Verónica, 1 0, 16-III-2007 (Exp. UPM leg.).
Bicyclus hewitsoni (Doumet, 1861)
Río Riaco, XII-2005 (Exp. UPM leg.); Río San Antonio, 11-III-2007 (Exp. UPM leg.); Poza Verónica, 16-III-2007 (Exp. UPM leg.).
Bicyclus sciathis (Hewitson, 1866)
Campamento UPM-Campamento Hormiga, 1 0, 18-III-2007 (Exp. UPM leg.).
Estimación de la diversidad
De la aplicación de los índices de diversidad alfa obtenemos los siguientes resultados:
ÍNDICE DE MARGALEF: DMg= 8,781321612.
Para valores inferiores a 2 se considera una comunidad poco diversa. Por el contrario son significativos valores superiores a 5, interpretados como comunidades de alta diversidad.
ÍNDICE DE SIMPSON: λ= 0,041333333.
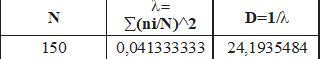
Como ya señalamos, en realidad se trata de un indicador de dominancia y, por tanto, se opone al de Diversidad. De este modo, es común hallar el complementario de l mediante la expresión DS=1- l, en nuestro caso resultando un valor de DS=0,95866667. No obstante es más frecuente el uso de la expresión opuesta a l, es decir, DS=1/ l. Mediante esta expresión, la Diversidad alcanza su mayor valor cuanto menor sea el valor de “lambda”, mientras la mínima diversidad viene dada por un índice de valor 1.
ÍNDICE DE SHANNON-WEAVER: H’ = 3,480556217.

Funciones de acumulación de especies
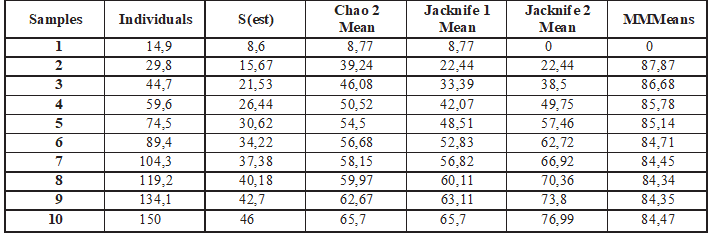
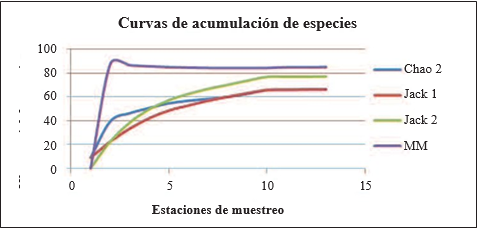
La aplicación de los modelos referidos en materiales y métodos arroja los siguientes resultados (Tabla 1) de las especies esperadas al aumentar el esfuerzo de los muestreos, es decir, las especies potencialmente presentes en la zona de estudio.
MODELO LINEAL
MM (Clench): Valor promedio 84,35 especies. Este modelo establece alrededor de 160 estaciones de muestreo para completar el inventario hasta alcanzar el 95% de las especies esperadas.
MODELOS NO PARAMÉTRICOS
Chao2: Arroja un valor del límite superior de 102,55 y un límite inferior de 52,44, con un promedio de 65,7 especies esperadas.
Jacknife 1: Se ajusta de manera exacta a los valores anteriores, promediando 65,7 especies potenciales.
Jacknife 2: Promedio 76,99 especies.
De la gráfica y valores anteriores basados en los modelos no paramétricos, se promedia una riqueza esperada de 69,46 especies de lepidópteros ninfálidos potencialmente presentes en la Caldera de Lu- bá. Con estos datos potenciales, las especies registradas en nuestros muestreos (45) elevan la tasa de capturas aproximadamente a un significativo 66% del total esperado.
Discusión
Siete especies de Nymphalidae mostradas en el presente trabajo, Amauris vashti, Bicyclus golo, Bicyclus neustetteri, Cymothoe althea, Cymothoe consanguis, Cymothoe capella y Lachnoptera anticlia representan las primeras citas para Bioko. De estas siete especies sólo Amauris aparece citada en la región continental (KHEIL, 1909), pero para ambos Bicyclus también suponen las primeras citas para Guinea Ecuatorial.
La UICN incluye, en la Lista Roja, a tres especies de las citadas en el presente trabajo, todas ellas bajo el epígrafe de Preocupación menor (LC) (LARSEN, 2011). Amauris echeria fernandina mantiene poblaciones estables en todo su área de distribución, si bien algunas subespecies pueden estar localmente amenazadas. Cymothoe caenis . Cymothoe consanguis son especies perfectamente adaptadas el bosque monzónico y, en conjunto, no están amenazadas aun cuando sus efectivos poblacionales suelen ser escasos (LARSEN, 2011).
De los resultados obtenidos tras la aplicación de los diferentes índices de diversidad, podemos inferir importantes conclusiones. Valores superiores a 5 en la aplicación del índice de riqueza de Margalef reflejan alta diversidad de especies (MAGURRAN, 1998) resultando en nuestro caso un valor aproximado de 8,8. Este indicador puede, por sí mismo, orientarnos acerca de la riqueza específica de los Nymphalidae de la Caldera de Lubá, al tiempo que nos revela una comunidad muy diversa. El cálculo de la lambda (λ) de Simpson arroja un valor muy bajo (0.041) y, como ya vimos, al tratarse de un indicador de dominancia la diversidad se halla precisamente por el valor inverso de λ, resultando por tanto una comunidad de gran diversidad (D = 24,19) (NÚÑEZ, 1991). En relación al índice de equidad de Shannon-Weaver (H’) MARGALEF (1972) demostró que los valores de este índice oscilan entre 1.5 y 3.5 y, en muy raras ocasiones, rebasan la cifra de 4.5. De esto podemos inferir que nuestros datos, con valores de H’ aproximadamente 3.5, denotan una estructura equitativa y una comunidad de muy alta diversidad. No obstante, se puede observar cierta desproporción en el número de capturas, con una mayor abundancia de unas pocas especies frente a otras. Este aparente desequilibrio es fácilmente explicable por la mayor capturabilidad de algunas especies (TURLIN, 2007).
Como acabamos de señalar, la zona de muestreo se enclava en un único tipo de hábitat lo cual, por sí mismo, puede justificar un número de especies relativamente bajo en relación al número de especies total de Nymphalidae de Bioko (SPEARMAN et al., 2000; Martín et al., en prep). No obstante, en el caso del presente trabajo la proporción hallada entre las especies inventariadas y las potenciales quedó establecida en un significativo 69%. Estos datos son parejos y coherentes con los índices de diversidad basados en la riqueza que acabamos de exponer (MAGURRAN, 1998; LONGINO et al., 2002).
En la actualidad no hay un método objetivo que se use como criterio para predecir cuándo un inventario ha alcanzado un número casi completo de especies (>95%) y, salvando aquellos muestreos en los que se logre el número de especies aportado por la asíntota de la curva (prácticamente inalcanzable en inventarios de invertebrados), solo se han establecido límites arbitrarios generalmente basados en proporciones entre los registros reales y los estimados por las curvas (JIMÉNEZ-VALVERDE & HOR- TAL, 2003; BECK & SCHWANGHART, 2010), o bien asumir como valor la pendiente de la curva en cada momento, es decir, la tasa de entrada de nuevas especies. En todo caso, para valores alrededor del 70% de las especies potencialmente presentes, las curvas de estimas en modelos no paramétricos se ha- cen estables y, por tanto, podemos considerar nuestro muestreo como representativo (MORENO & HALFFTER, 2001; JIMÉNEZ-VALVERDE et al., 2006; WILLIAMS, 2008).
Actualmente se estima que la familia Nymphalidae cuenta con alrededor de 140 especies en Bioko (Martín et al., en prep.) recogiendo, obviamente, todos los ambientes presentes en la isla, incluidos los urbanos, agrícolas, bosques degradados, etc. Nuestros registros, obtenidos exclusivamente en hábitat de pluvisilva monzónica representan alrededor del 33% de las especies de citadas para el conjunto de Bioko, aun cuando la superficie muestreada ha significado escasamente un 1,3% de la superficie de la Isla. El sur de Bioko permanece prácticamente inexplorado (MARTÍN & COBOS, 2010; NAVARRO et al., 2012), por lo que nuevos trabajos de muestreo podrán, sin duda, aportar nuevas especies. Obviamente si aumenta la muestra -otros trabajos de recolección- los índices variarán su valor pues todos ellos tien den a aumentar con el tamaño de la muestra (MORENO, 2001; GONZÁLEZ-OREJA et al., 2010; MO- RENO et al., 2011).

Reserva Científica de la Gran Caldera: 1. Fondo Caldera: 3º 31’ N 8º 31’ E (1.293 msnm), 2. Camp Ureca-Fondo Caldera 3º 27’ N 8º 31’ E (1.066 msnm), 3. Campamento Ureka: 3º 21’ 34’’ N 8º 30’ E (916 msnm), 4. Río San Antoni/Poza Verónica 3º 20’ N 8º 29’ E (899 msnm), 5. Pizarras: 3º 21’ N 8º 29’ E (875 msnm). 2. Zonas de captura, 6. Camp. UPM y Río Riaco: 3º 20’ N 8º 29’ E (876 msnm), 7. Camp. UPM/Camp. Hormiga 3º 19’ N 8º 28’E (713 msnm), 8. Campamento Hormiga: 3º 18’ N 8º 28’ E (585 msnm), 9. Chapa Herminio: 3º 17’ N 8º 27’ E (374 msnm), 10. Moraca: 3º 15’ N 8º 28’ E (nivel mar). 3. Amauris vasthi (Butler, 1869) reverso. 4. Amauris vasthi (Butler, 1869) anverso. 5. Bicyclus golo (Aurivillius, 1893) macho reverso. 6. Bicyclus golo (Aurivillius, 1893) macho anverso.

Bicyclus neustetteri (Rebel, 1914) macho anverso. 8. Bicyclus neustetteri (Rebel, 1914) hembra reverso. 9.Acraea abdera abdera Hewitson, 1852, macho. 10. Cymothoe oemilius fernandina Hall, 1929, macho. 11. Kallimoides rumia (Doubleday, 1849) macho. 12. Amauris inferna moka Talbot, 1940.
Agradecimientos
Queremos mostrar muestro especial agradecimiento a Pablo Cobos y Pedro Paniagua por su ayuda en la recolección y preparación de los ejemplares. Asimismo a Ignacio Arizmendi por su activa colaboración en la elaboración de las curvas de acumulación, lo que sin duda ha supuesto un notable aporte a este trabajo. Ambas Expediciones de la Universidad Politécnica de Madrid realizadas en 2005 y 2007 a la Caldera de Lubá, se realizaron con el patrocinio del Programa Nacional de I+D+I (CGL2005-23762- E y CGL2006-27110-E/BOS).
BIBLIOGRAFÍA
ACKERY, P. R., SMITH, C. R. & VANE-WRIGHT, R. I., 1995.– Carcasson’s African Butterflies: An annotated Ca- talogue of the Papilionoidea and Hesperioidea of the Afrotropical Region : 803 pp. British Museum (Natural History). London.
ACKERY, P. R., DE JONG, R. & VANE-WRIGHT, R. I., 1999.– The butterflies: Hedyloidea, Hesperioidea and Pa- pilionoidea.– In N. P. KRISTENSEN (ed.). Handbook of Zoology, .(35): 263-300. De Gruyter, Berlin.
BAEV, P. V. & PENEV, L. D., 1995.– BIODIV: program for calculating biological diversity parameters, similarity, niche overlap, and cluster analysis. Version 5.1.: 57 pp. Pensoft, Sofia-Moscow.
BECK, J. & SCHWANGHART, W., 2010.– Comparing measures of species diversity from incomplete inventories: an update.– Methods in Ecology & Evolution, .: 38-44.
BONEBRAKE T., PONISIO, C., BOGGS, C. L. & EHRLICH, P. R., 2010.– More than just indicators: A review of tropical butterfly ecology and conservation.– Biological Conservation, 143: 1831-1841.
BROWER, A. V. Z., 2000.– Phylogenetic relationships among the Nymphalidae (Lepidoptera), inferred from partial sequences of the wingless gene.– Proceedings of the Royal Society of London (B), 267: 1201-1211.
BURGESS, N., HALES, J. D., RICKETTS, T. H. & DINERSTEIS, E., 2006.– Factoring species, non-species values and threats into biodiversity prioritisation across the ecoregions of Africa and its islands.– Biological Conservation, 127: 383-401.
BUTYNSKI, T. M. & KOSTER, S. H., 1994.– Distribution and conservation status of primates in Bioko Island, Equatorial Guinea.– Biodiversity and Conservation, .: 893-909.
CHAO, A., 1984.– Non–parametric estimation of the number of classes in a population. Scandinavian Journal of Statisics, 11: 265-270.
CHAO, A., 2005.– Species richness estimation.– In N. BALAKRISHNAN, C. B. READ & B. VIDAKOVIC Eds. Encyclopedia of Statistical Sciences: 7909-7916. Wiley, New York.
COLWELL, R. K., 2006.– EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples. Version 9. User’s Guide and application published. Available from http://purl.oclc.org/estimates. (Accedido el 3 de octubre de 2014).
COLWELL, R. K., MAO, C. X. & CHANG, J., 2004.– Interpolating, extrapolating, and comparing incidence- based species accumulation curves.– Ecology, 85: 2717-2727.
ESPINOSA, T., 2003.– ¿Cuántas especies hay? Los estimadores no paramétricos de Chao.– Elementos de ciencia y cultura, 52: 53-56.
FREITAS, A. V. L. & BROWN, K. S., 2004.– Phylogeny of the Nymphalidae (Lepidoptera).– Systematic Biology,
COLWELL, R. K., 2006.– EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples. Version 9. User’s Guide and application published. Available from http://purl.oclc.org/estimates. (Accedido el 3 de octubre de 2014).
COLWELL, R. K., MAO, C. X. & CHANG, J., 2004.– Interpolating, extrapolating, and comparing incidence- based species accumulation curves.– Ecology, 85: 2717-2727.
ESPINOSA, T., 2003.– ¿Cuántas especies hay? Los estimadores no paramétricos de Chao.– Elementos de ciencia y cultura, 52: 53-56.
FREITAS, A. V. L. & BROWN, K. S., 2004.– Phylogeny of the Nymphalidae (Lepidoptera).– Systematic Biology, 53: 363-383.
FUSTER J. M., 1956.– Un accidente volcánico excepcional: la Caldera de San Carlos (Fernando Poo).– Archivos IDEA, 40: 65-74.
GOTELLI, N. J. & COLWELL, R. K., 2001.– Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness.– Ecology Letters, 4: 379-391.
GONZÁLEZ-OREJA, J. A., DE LA FUENTE, A. A., HERNÁNDEZ, L., BUZO, D. & BONACHE, C., 2010.– Evaluación de estimadores no paramétricos de la riqueza de especies. Un ejemplo con aves en áreas verdes de la ciudad de Puebla, México.– Animal Biodiversity and Conservation, 33(1): 31-45.
JIMÉNEZ-VALVERDE, A. & HORTAL, J., 2003.– Las curvas de acumulación de especies y la necesidad de eva- luar la calidad de los inventarios biológicos.– Revista Ibérica de Aracnología, 8: 151-161.
HALFFTER, G. & EZCURRA, E., 1991.– ¿Qué es la Biodiversidad?.– In G. HALFFTER. La Diversidad Biológica de Iberoamérica: 3-24. Acta Zoológica Mexicana (n.s.): 389 pp. CYTED-D, Programa Iberoamericano de Ciencia y Tecnología para el Desarrollo, México D. F.
HARVEY, D. J., 1991.– Higher classification of the Nymphalidae.– In H. F. NIJHOUT. The Development and Evo- lution of Butterfly Wing Pattems: 255-273. Smithsonian lnstitution Press, Washington D. C.
HAYWOOD, V. H., 1994.– The measurement of biodiversity and the politics of implementation.– In P. L. FOREY, C. J. HUMPHRIES & R. I. VANE-WRIGHT (Eds). Systematic and Conservation Evaluation, 15-22: 438 pp. Oxford.
HENNING, S., 1988.– The Charaxinae butterflies of Africa: 457 pp. Ed. Frandsen. Horsens.
JIMÉNEZ-VLAVERDE, A. & HORTAL, J. 2003.– Las curvas de acumulación de especies y la necesidad de evaluar la calidad de los inventarios biológicos.– Revista Ibérica de Aracnología, 8: 151-161.
JIMÉNEZ-VALVERDE, A., JIMÉNEZ-MENDOZA, S., MARTÍN-CANO, J. & MUNGUIRA, M. L., 2006.– Comparing relative model fit of several species–accumulation functions to local Papilionoidea and Hesperioidea butterfly inventories of Mediterranean habitats.– Biodiversity and Conservation, 15: 177-190.
JOST, L., 2010.– The relation between evenness and diversity.– Diversity, 2: 207-232.
KHEIL, N. M., 1909.– Catálogo sistemático de la fauna de las posesiones españolas del Golfo de Guinea (Lepidóp- teros).– Memorias de la Sociedad Española de Historia Natural I, Memoria 1(28): 483-506.
LARSEN, T. B., 2005.– Butterflies of West Africa: 595pp. Apollo Books, Svendborg.
LARSEN, T. B., 2011.– The IUCN Red List of Threatened Species. Version 2014.2. Available from http://www.iucn- redlist.org. (accessed 16 September 2014).
LONGINO, J. T., CODDINGTON, J. & COLWELL, R. K., 2002.– The ant fauna of a tropical rain forest: estimating species richness three different ways.– Ecology, 83: 689-702.
MADDISON, D. R. & SCHULZ, K. S., 2007.– The Tree of Life Web Project. Available from http://tolweb.org. (ac- cessed 25 April 2014).
MARGALEF, R., 1958.– Information theory in ecology.– General Systematics, 3: 36-71.
MARGALEF, R., 1972.– Homage to Evelyn Hutchinson, or why there is an upper limit to diversity.– Transactions of the Connecticut Academy of Arts and Sciences, 44: 211-235.
MAGURRAN, A. E., 1998.– Ecological diversity and its measurement:. 179 pp. Princeton University Press, New Jersey.
MAGURRAN, A. E., 2007.– Species abundance distributions over time.– Ecology Letters, 10: 347-354.
MARTÍN, I. & COBOS, P., 2010.– Expedición Científica a la Caldera de Lubá. Isla de Bioko (Guinea Ecuatorial).– In E. VIGUERA, A. GRANDE & J. LOZANO (Coordinadores). Encuentros con la Ciencia II. Del macrocos- mos al microcosmos: 137-150. Servicio de Publicaciones de la Universidad de Málaga. Málaga.
MARTÍN, I., ARREDONDO, A. & VÉLEZ, Y., 2014.– Anfibios y reptiles del Parque Nacional de las Tablas de Daimiel: 167 pp. Organismo Autónomo de Parques Nacionales. Madrid.
MARTÍNEZ, T., 1968.– Fernando Poo: geografía, historia, paisaje: 119 pp. La Guinea Española Ediciones, Instituto Claretiano Africanista Santa Isabel.
MORENO, C. E., 2001.– Métodos para medir la biodiversidad.– Manuales y Tesis Sociedad Entomológica Aragone- sa, 1: 1-84.
MORENO, C. E. & HALFTER, G., 2001.– On the measure of sampling effort used in species accumulation cur- ves.– Journal of Applied Ecology, 38: 487-490
MORENO, C. E., BARRAGÁN, F., PINEDA, E. & PAVÓN, N., 2011.– Reanálisis de la diversidad alfa: alternativas para interpretar y comparar información sobre comunidades ecológicas.– Revista Mexicana de Biodiversidad, 82: 1249-1261.
NAVARRO, R. M., CLEMENTE, M., KASIMIS, N., PADRÓN, E., HERNÁNDEZ, E., MARTÍN, E. & GARCÍA, A., 2012.– Cartografía de la vegetación de la Isla de Bioko (Guinea Ecuatorial) mediante el uso de imágenes LandSat 7 ETM+: particularización del piso afromontano.– Darwiniana, 50(2): 252-265.
NÚÑEZ, E., 1991.– Sobre la cuantificación de la diversidad ecológica.– Hidrobiológica, 1(1): 87-93.
NYLIN, S. SLOVE, J. & JANZ, N., 2014.– Host plant utilization, host range oscillations and diversification in nymphalid butterflies: a phylogenetic investigation Evolution.– International Journal of Organic Evolution, 68(1): 105-124.
OBAMA, C., 2006.– De la expansión a la recesión: la inoperancia legal en la gestión de los recursos forestales de Guinea Ecuatorial: 166-181. Movimiento Mundial por los Bosques Tropicales, WRN.
PALMER, M. W., 1990.– The estimation of species richness by extrapolation.– Ecology, 71: 1195-1198.
TURLIN, B., 1999.– Observations sur les espèces insulaires africaines de la sous-famille des Charaxinae (Lepidop- tera: Nymphalidae).– Lambillionea, 49: 171-182.
TURLIN, B., 2007.– Butterflies of the World, Part 28: Nymphalidae, No. 14: The Afrotropical Species of Charaxes, Part 3: 40 pp. Antiquariat Goecke & Evers, Keltern.
RICKETTS, T. H.; DAILY, G. C. & EHRLICH, P. R., 2002.– Does butterfly diversity predict moth diversity? Tes- ting a popular indicator taxon at local scales.– Biological Conservation, 103: 361-370.
RICOTTA, C., 2005.– Through the jungle of biological diversity.– Acta Biotheoretica, 53: 29-38.
SÁFIÁN S. Z., COLLINS, S. C., KORMOS, B. & SIKLÓSI, A., 2009.– African Butterfly Database version 1.0. Available from http://www.abdb-africa.org. (accessed 7 October 2014).
SAMWAYS, M. J., 1994.– Insect Conservation Biology: 358 pp. Chapman & Hall, London.
SOBERÓN, J. & LLORENTE, J., 1993.– The use os species accumulation functions for the prediction of species richness.– Conservation Biology, 7: 480-488.
SPEARMAN, L. A., ORFE, N. A. & WEINTRAUB, J. D., 2000.– An annotated list of the butterfly fauna of Bioko Island, Equatorial Guinea (Lepidoptera: Papilionoidea, Hesperioidea).– Transactions of the American Entomo- logical Society, 126(3-4): 447-475.
VANE-WRIGHT R, HUMPHRIES, C. & WILLIAMS, P. 1991.– What to protect? Systematics and the agony of choice. Biological Conservation 55: 235-254.
VANE-WRIGHT, R. I., 2003.– Evidence and identity in butterfly systematics.– In C. L. BOGGS, W. B. WATT & P. R. EHRLICH (eds). Butterflies: Ecology and Evolution Taking Flight: 477-513. University of Chicago Press, Chicago.
WAHLBERG, N., WEINGARTNER, E. & NYLIN, S., 2003.– Towards a better understanding of the higher syste- matics of Nymphalidae (Lepidoptera: Papilionoidea).– Molecular Phylogenetics and Evololution, 28: 473-484.
WAHLBERG, N., BRABY, M. F., BROWER, A. V. Z., DE JONG, R., LEE, M. M., NYLIN, S., PIERCE, N., ES- PERLING, F., VILA, R., WARREN, A. D. & ZAKHAROV, E., 2005.– Synergistic effects of combining morp- hological and molecular data in resolving the phylogeny of butterflies and skippers. Proceedings of the Royal Society Series B 272:1577-1586.
WAHLBERG, N., LENEVEU, J., KODANDARAMAIAH, U., PEÑA, C., NYLIN, S., FREITAS, A. V. L. & BROWER, A. V. Z., 2009.– Nymphalid butterflies diversity following near demise at the Cretaceous/Tertiary boun- dary.– Proceedings of the Royal Society Series B Biological Sciences, 276: 4295-4302.
WHITTAKER, R. H., 1972.– Evolution and measurement of species diversity.– Taxon, 21: 213-251.
WILLIAMS, M. C., 2008.– Checklist of Afrotropical Papilionoidea and Hesperoidea.– Compiled by Mark C. Wi- lliams, 7th ed. Available from http://www.atbutterflies.com/index.htm (accessed 18 August 2014).
WILLIAMS, M. R., 2008.– Assessing diversity of diurnal Lepidoptera in habitat fragments: testing the efficiency of strip transects. -Environmental Entomology, 37: 1313-1322.
ZAMORA, C.; PARRA, E. & JAQUE, E., 2011.– Patrones de distribución de los geométridos de la Región del Bio- bío, Chile: Una aproximación para su conservación.– Revista Chilena de Historia Natural, 84(4): 465-480.
ZHANG, M., CAO, T., JIN, K., REN, Z., GUO, Y., SHI, J., ZHONG, Y. & MA, E., 2008.– Estimate divergence times among subfamilies in Nymphalidae.– Chinese Science Bulletin, 53(17): 2652-2658.