
Contribución al conocimiento de los Geometridae Leach, 1815 del Parque Natural Cabo de Gata-Níjar (Almería, España) (Lepidoptera: Geometridae)
Contribution to the knowledge of the Geometridae Leach, 1815 in the Cabo de Gata-Nijar (Almeria, Spain) Natural Park (Lepidoptera: Geometridae)
Contribución al conocimiento de los Geometridae Leach, 1815 del Parque Natural Cabo de Gata-Níjar (Almería, España) (Lepidoptera: Geometridae)
SHILAP Revista de lepidopterología, vol. 44, núm. 173, 2016
Sociedad Hispano-Luso-Americana de Lepidopterología
Recepción: 05 Junio 2014
Aprobación: Julio , 21, 2014
Resumen: La familia Geometridae está representada en el Parque Natural Cabo de Gata-Níjar (Almería, sureste de España) por 104 especies pertenecientes a las subfamilias Ennominae, Desmobathrinae, Geometrinae, Sterrhinae y Larentiinae. El corotipo atlanto-mediterráneo es ampliamente mayoritario (58,7%), incluyendo 8 endemismos ibéricos. Del total de especies, 20 son nuevas para la provincia de Almería y 48 para el territorio del parque natural y su entorno.
Palabras clave: Lepidoptera, Geometridae, Parque Natural Cabo de Gata-Níjar, Almería, España.
Abstract: The Geometridae fauna in the Cabo de Gata-Níjar (Almeria, southern Spain) Natural Park includes 104 species belonging to the subfamilies Ennominae, Desmobathrinae, Geometrinae, Sterrhinae and Larentiinae. The atlantomediterranean corotype is widely represented (58.7%), including 8 Iberian endemisms. The study contributes with twenty new records for Almeria province and forty-eight new records for the park and surrounding area.
Keywords: Lepidoptera, Geometridae, Cabo de Gata-Nijar Natural Park, Almeria, Spain.
Introducción
El Parque Natural Cabo de Gata-Níjar es un área de especial interés entomológico debido a su situación geográfica en la península ibérica y por la proximidad con el norte de África, destacando sus excepcionales valores paisajísticos, geológicos y botánicos y sus particulares características climáticas que lo convierten en el punto más seco y cálido de Europa. Sin embargo, el conocimiento de su fauna lepidopterológica y de la familia Geometridae en particular, parece claramente insuficiente. El mayor volumen de capturas es aportado por KRAUS (1997, 1999), que cita 29 especies en unos muestreos realizados en primavera y otoño; otras resultan de las visitas esporádicas al parque realizadas por AGENJO (1952), LAJONQUIERE (1963), HACKMAN (1968), HACKER & WOLF (1982), MUÑOZ (1992) y ORTIZ et al. (2010) coincidiendo, como es habitual, con los meses más favorables del año. Además, otras citas se pueden consultar en obras de carácter general (HAUSSMAN, 2001, 2004; REDONDO et al., 2009; HAUSMANN & VIIDALEPP, 2012).
El Parque Natural Cabo de Gata-Níjar está situado en el sureste de la península ibérica compartiendo parte de los municipios de Almería, Carboneras y Níjar. El área de estudio ocupa una superficie de 37.500 ha terrestres y está formado por cuatro unidades geomorfológicas: la sierra de Cabrera, la depresión de Níjar, la sierra de Cabo de Gata y la llanura litoral de la Bahía de Almería (Figura 1).
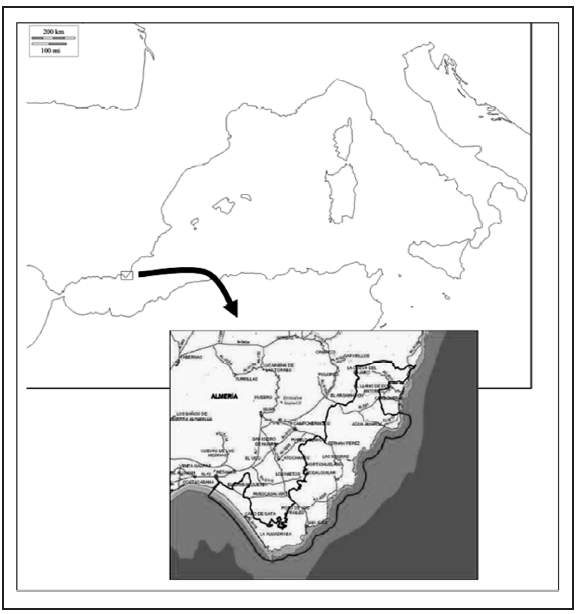
Geológicamente, este espacio natural forma parte del sector suroriental de la Cordillera Bética, limitando al norte con las estribaciones de la Sierra de Cabrera, que está formada por sustratos carbonatados y silicatados y cuya mayor altitud es La Serrata (562 m). Al este, y prolongándose como una franja litoral de relieve muy accidentado, se alza la sierra de Cabo de Gata y Carboneras, constituida por rocas volcánicas ácidas y con el Cerro del Fraile (492 m) como mayor elevación. Al pie de estos macizos montañosos se han depositado los sedimentos cuaternarios de origen erosivo que rellenan las llanuras interiores (depresión de Níjar). Por otra parte, las llanuras litorales comprenden las playas, dunas y saladares, cuya mayor expresión se da al suroeste del territorio (dunas de Torregarcía, salinas de Cabo de Gata).
Bioclimáticamente, el área de estudio se define como xérico-oceánico, con temperaturas medias anuales entre 18 y 20º C y precipitaciones medias anuales entre 180 y 270 mm. Se puede distinguir el piso bioclimático inframediterráneo limitado a la estrecha franja terrestre más cercana al mar, y el piso termomediterráneo en el resto del territorio. Los ombrotipos presentes son el árido y el semiárido.
Biogeográficamente, el Parque Natural se encuadra en la provincia corológica Murciano- Almeriense, sector Almeriense y subsector Caridemo (PEINADO et al., 1992). Las series climatófilas reconocidas en el área de estudio son la serie del cornical (Mayteno europaei-Periploco angustifoliae S.), extendida en los pisos infra y termomediterráno inferior, con ombrotipos árido y semiárido y localizada en solanas abruptas, donde se ve favorecida por el efecto amortiguador de los vientos marinos húmedos. Cuando se incrementa la continentalidad y la aridez, aquella es sustituida por la serie termomediterránea superior, semiárida del lentisco (Chamaeropo humilis- Rhamno lycioidis S.). En las zonas llanas y sobre materiales sedimentarios se presenta la serie infratermomediterránea, árido-semiárida del azufaifo (Zizipho loti S.). En la actualidad, la mayor parte de la superficie que podrían ocupar las comunidades climácicas está colonizada por sus etapas seriales (retamares, espartales, tomillares, matorrales nitrófilos, etc.) y por cultivos de secano o regadío.
En aquellos ambientes donde las características edáficas o microtopográficas son condicionantes, se presentan diversas comunidades permanentes como los retamares edafoxerófilos, que colonizan amplias extensiones de roquedos y pedregales en áreas montañosas, al contrario que las comunidades temporihigrófilas que prosperan muy discretamente en los cauces fluviales secos y ramblas que surcan el territorio. En las llanuras litorales, ya sea por efecto de la arena, la maresía y el viento, o de la salinidad y humedad del suelo, se reconocen las geopermaseries de playas y dunas litorales (psammófilas) y de saladares (halófilas), respectivamente.
El objetivo de este trabajo es ofrecer un catálogo sistemático de las especies de la familia Geometridae presentes en el Parque Natural Cabo de Gata-Níjar, realizar su análisis biogeográfico y fenológico y destacar las especies más interesantes.
Material y métodos
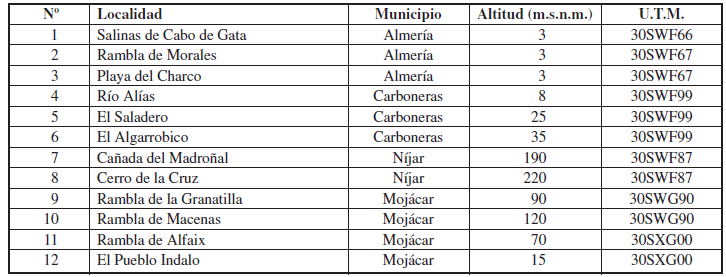
Se presentan los resultados de los 54 muestreos nocturnos realizados en varias localidades del Parque Natural Cabo de Gata-Níjar, a los que se suma la localidad limítrofe de la Rambla de la Granatilla, durante el período comprendido entre la segunda quincena de mayo de 2012 y la primera quincena de mayo de 2014. Además, se han considerado algunos otros muestreos esporádicos efectuados en años anteriores en parajes colindantes del municipio de Mojácar. La totalidad de las localidades estudiadas se ordenan por municipios en la Tabla 1, aunque los muestreos periódicos se han realizado en las nueve primeras. Estas localidades tienen las siguientes características:
Estaciones 1-3. Se localizan en el ámbito de los arenales y saladares litorales, colonizados por una flora con apetencias ecológicas estrictas y adaptadas a medios muy especializados. Estaciones 4-6. Se ubican en el dominio de la serie fitosociológica del cornical, encontrándose la comunidad cabeza de serie, en general, muy bien conservada y con una alta diversidad florística. Estaciones 7- 8. Se sitúan en el dominio de la serie del lentisco, representada principalmente por sus etapas seriales (espartales y tomillares). Estación 9. Se halla en el entorno de los retamares edafoxerófilos.
Los muestreos se han realizado utilizando trampas de luz negra y actínica de 6 vatios (tipo Heath).
La relación de especies estudiadas se puede consultar en el Apéndice, ordenadas sistemáticamente e indicando, para cada taxón, la toponimia distintiva, fecha de captura u observación, número de ejemplares, corotipo, fenología y referencias bibliográficas. Las especies que se citan por vez primera para la provincia de Almería están marcadas con un asterisco (*) y con dos asteriscos (**) para el P.N. Cabo de Gata-Níjar. Asimismo, se indica el período de vuelo (en meses y numeración romana) que se ha confirmado en base a las capturas, observaciones y referencias bibliográficas específicas. En lo que respecta a las citas bibliográficas, se han seleccionado aquellas referidas expresamente al ámbito del parque natural, aunque también se han tenido en consideración algunas localidades limítrofes (Sierra de Cabrera, Retamar y El Alquián).
La nomenclatura y la ordenación de los taxones en sus correspondientes categorías taxonómicas se han hecho de acuerdo con la propuesta de Fauna Europaea (2013) y REDONDO et al. (2009). Los datos biogeográficos y fenológicos se han obtenido mayoritariamente de las principales obras de carácter generalista (HAUSMAN, 2001, 2004; MIRONOV, 2003; ROBINEAU, 2007; REDONDO et al., 2009, LERAUT, 2009; HAUSMANN & VIIDALEPP, 2012).
Resultados y discusión
La fauna de Geometridae del Parque Natural Cabo de Gata-Níjar está compuesta, hasta el momento, por 104 especies, distribuidas en las subfamilias Ennominae (27 especies), Desmobathrinae (1), Geometrinae (7), Sterrhinae (39) y Larentiinae (30). De éstas, 20 son nuevas para la provincia de Almería y 48 lo son para el territorio del Parque Natural y su entorno más inmediato.
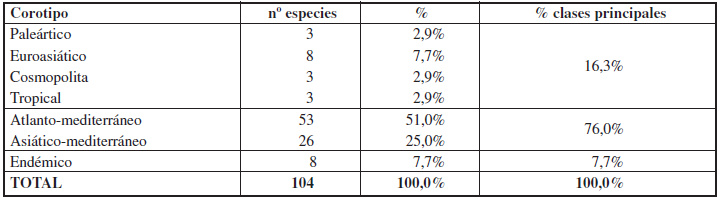
Biogeográficamente, la influencia mediterránea, sumando los endemismos, es ampliamente mayoritaria (83,7%), en correspondencia con la posición geográfica del área de estudio en el continente europeo, destacando los elementos atlanto-mediterráneos (51%) sobre los asiáticomediterráneos (25,0%) (Tabla 2). Los taxones endémicos de la península ibérica representan el 7,7% del total con 8 especies. El importantísimo número de especies que se distribuyen exclusivamente por Europa occidental y el noroeste de África, evidencia la condición de este espacio protegido como refugio para una fauna que vive en un área geográfica reducida. Entre los elementos de amplia distribución destacan ligeramente los euroasiáticos (7,7%).
En la subfamilia Ennominae destacan los endemismos Menophra annegreteae Skou, 2007, descrita de Almería, Sardocyrnia fortunaria (Vázquez, 1905) y Charissa assoi (Redondo & Gastón, 1997). Esta última especie vive en una hábitat diferenciado geográfica, climática y ecológicamente de las localidades en las que anteriormente se conocía su presencia (GARRE et al., 2013). Entre las especies de esta subfamilia, la captura de la especie ibero-magrebí Lhommeia biskraria (Oberthür, 1885) confirma su presencia en el sureste de la península ibérica junto con las citas en las localidades almerienses de La Garrofa (REDONDO et al., 2009) y El Alquián (MAGRO, 2013).
La subfamilia Desmobathrinae está representada por Myinodes interpunctaria (Herrich- Schäffer, 1939), especie escasamente citada en la mitad sur peninsular (REDONDO et al., 2009).
La subfamilia Geometrinae incluye el elemento paleotropical Pingasa lahayei (Oberthür, 1887), solamente conocido en Europa de las localidades almerienses de Cabo de Gata (VALLHONRAT, 1980; KRAUS, 1997, 1999) y Villaricos (REDONDO et al., 2009).
En la subfamilia Sterrhinae destaca la presencia de cinco especies endémicas: Idaea alicantaria (Reisser, 1963), I. urcinata (Agenjo, 1952), solamente conocida de Almería, I. lutulentaria (Staudinger, 1892), I. deitanaria Reisser & Weisert, 1977 y Brachyglossina hispanaria (Püngeler, 1913). La presencia de las especies simpátricas y sincrónicas Idaea alyssumata (Millière, 1871) e I. urcinata, implica el estudio de sus genitalias para evitar errores de identificación, aunque el color más claro del fondo de las alas y los puntos discales más contrastados en I. urcinata son caracteres muy constantes y útiles para diferenciar ambas especies (HAUSSMAN, 2004). Otros taxones interesantes que tienen una distribución típicamente meridional en la Península Ibérica son Idaea attenuaria (Rambur, 1833), I. rainerii Hausmann, 1994, I. blaesii Lenz & Hausmann, 1992, I. fractilineata (Zeller, 1847) y Oar reaumuraria (Millière, 1864) (REDONDO et al., 2009).
La familia Larentiinae tiene entre sus representantes más destacados a Epirrhoe sandosaria (Herrich-Schäffer, 1852), Larentia malvata (Rambur, 1833), Horisme scorteata (Staudinger, 1901), Eupithecia orana Dietze, 1910 y E. unitaria Herrich-Schäffer, 1851. Un caso singular sería la captura de un ejemplar de Epirrita dilutata ([Denis & Schiffermüller], 1775) de la que se tienen escasos datos en la mitad sur peninsular (REDONDO et al., 2009; HAUSMANN & VIIDALEPP, 2012). La especie se capturó en un hábitat extremadamente xerofítico, cálido y prácticamente desprovisto de arbolado que contrasta con sus requerimientos mesófilos y silvícolas (REDONDO et al., 2009; HAUSMANN & VIIDALEPP, 2012). En cuanto a la posible planta nutricia de la oruga, de acuerdo con HAUSMANN & VIIDALEPP (2012), solamente se ha constatado la presencia, a relativa distancia, de algunos bosquetes autóctonos de álamo blanco (Populus alba).
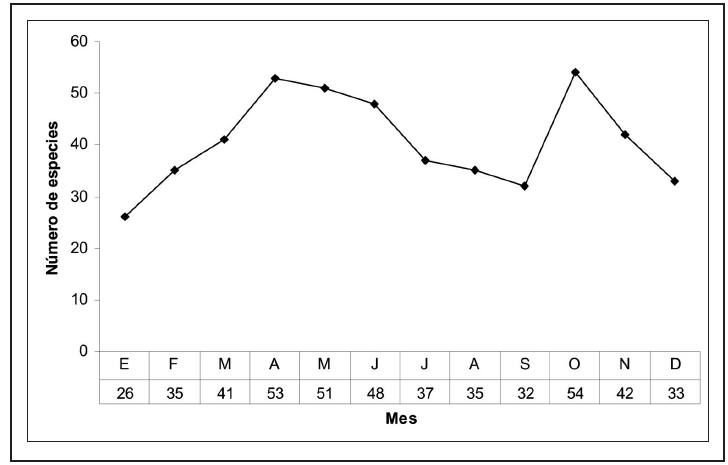
Fenológicamente, la abundancia relativa de especies durante todos los meses del año varía entre 26 y 54 especies y parece estar relacionada con las condiciones climáticas anuales del área de estudio, ya que los inviernos son muy suaves y similares a la primavera de latitudes superiores. De esta manera, el mayor número de especies se concentra en primavera y otoño, con su máximo en abril y octubre, disminuyendo moderadamente durante los meses más fríos desde diciembre a febrero y en la diapausa estival, debido a las condiciones de aridez extrema y a un agostamiento generalizado de la vegetación (Figura 2). Al margen de la subfamilia Desmobathrinae, que solo cuenta con un único representante de vuelo invernal, las subfamilias Ennominae y Larentiinae se ajustan al esquema fenológico general, mientras que las subfamilias Geometrinae y Sterrhinae desplazan sus máximos en el final de la primavera y al comienzo del verano.
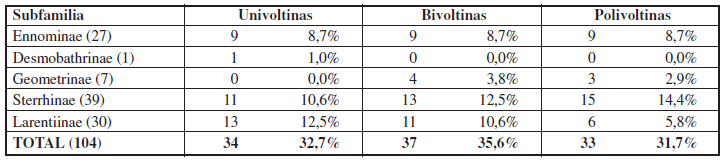
En relación al ciclo vital, las especies univoltinas suponen el 32,7% del total, las bivoltinas el 35,6% y las polivoltinas el 31,7% (Tabla 3), lo que parece estar relacionado con las condiciones climáticas y la disponibilidad de alimento durante la mayor parte del año, que permite a las especies reproducirse en sucesivas generaciones.
La fenología de los imagos confirmada en este territorio parece coincidir con la información extraída de las fuentes bibliográficas, muy especialmente en el caso de las especies univoltinas. Sin embargo, el período de vuelo en las especies bivoltinas y polivoltinas se mantiene en muchos casos sin interrupciones, solapándose las sucesivas generaciones. En tales circunstancias, se ha renunciado a delimitar los intervalos generacionales y se ha optado por indicar el periodo de vuelo completo en el Apéndice.
En el caso de los taxones que son, aparentemente, muy escasos en el territorio y de los cuales se cuenta con muy pocos registros parece que su período de vuelo no tiene que coincidir necesariamente con el observado.
Agradecimientos
Queremos agradecer a la Dirección del Parque Natural Cabo de Gata-Níjar las facilidades prestadas para el desarrollo de este estudio.
BIBLIOGRAFÍA
AGENJO, R., 1952.– Fáunula lepidopterológica Almeriense: 370 pp., 24 pl. CSIC, Madrid.
AGENJO, R., 1964.– Sección de capturas IV.– Graellsia, 20: 191-202. EXPÓSITO-HERMOSA, A., 1985.– Apuntes sobre la familia Geometridae (sensu stricto).– SHILAP Revista de lepidopterología, 13(49): 27-31.
FAUNA EUROPAEA, 2013.– Fauna Europaea ver. 2.6. Disponible en http://www.faunaeur.org (accedido el 6 de abril de 2014).
GARRE, M., RUBIO, R. M., GUERRERO, J. J. & ORTIZ, A. S., 2013.– Nuevos datos sobre la distribución de Charissa (Euchrognophos) assoi (Redondo & Gastón, 1997) en la mitad sur de la Península Ibérica (Lepidoptera: Geometridae, Ennominae).– Boletín de la Sociedad Andaluza de Entomología, 21: 22-27.
HACKER, H. & WOLF, W., 1982.– Contribution to the Lepidoptera-fauna of Spain. II. Heterocera of a threeweek visit in summer 1980 (First part).– SHILAP Revista de lepidopterología, 10(40): 267-273.
HACKMAN, W., 1968.– Lepidoptera of Southern Spain.– Notulae Entomologicae, 48: 149-168.
HAUSMANN, A., 2001.– Introduction. Archierinae, Orthostixinae, Desmobathrinae, Alsophilinae, Geometrinae.– In A. HAUSMANN (Ed.). The Geometrid moths of Europe, 1: 282 pp. Apollo Books, Stenstrup.
HAUSMANN, A., 2004.– Sterrhinae.– In A. HAUSMANN (Ed.). The Geometrid moths of Europe, 2: 600 pp. Apollo Books, Stenstrup.
HAUSMANN, A. & VIIDALEPP, J., 2012.– Larentiinae I.– In A. HAUSMANN (Ed.). The Geometrid Moths of Europe, 3: 743 pp. Apollo Books, Stenstrup.
KRAUS, W., 1997.– A contribution to knowledge of the Lepidoptera of the “Parque Natural Cabo de Gata”, Nijar, Almeria, Spain.– SHILAP Revista de lepidopterología, 25(97): 63-64.
KRAUS, W., 1999.– Beobachtungen zur Macrolepidopterenfauna der Iberische Halbilsen. Teil. 2: Artenliste Drepanidae bis Notodontidae.– Nachrichten des Entomologischen Vereins Apollo, N.F., 20 (2): 231-263.
LAJONQUIERE, Y., 1963.– Captura en Almería de un Cossidae nuevo para la fauna europea (Lepidoptera).– Graellsia, 20: 29-31.
LERAUT, P., 2009.– Moths of Europe. Geometrid Moths, 2: 804 pp. N. A. P. Editions, Verrières-le-Buisson.
MAGRO, R., 2013.– Los lepidópteros heteróceros de la rambla del Puente de la Quebrada y Cueva del Meadero (Almería, España) y algunos casos de foresia detectados sobre Agrotis boetica (Boisduval, [1837]), Agrotis lasserrei (Bugnion, 1837) y Eremochlaena orana (Lucas, 1894) por el pseudoescorpión Diplotemnus insolitus Chamberlin, 1933. (Lepidoptera: Noctuidae. Pseudoscorpiones: Atemnidae).– Arquivos entomolóxicos, 8: 33-46.
MIRONOV, V., 2003.– Larentiinae II (Perizomini and Eupitheciini).– In A. HAUSMANN (Ed.). The Geometrid Moths of Europe, 4: 463 pp. Apollo Books, Stenstrup.
MUÑOZ, J., 1992.– Algunes troballes de lepidòpters fetes a la província d’Almeria.– Butlletí de la Societat Catalana de Lepidopterologia, 70: 28-29.
ORTIZ, A. S., GARRE, M., GUERRERO, J. J., RUBIO, R. M. & CALLE, J. A., 2010.– Nuevos datos sobre los macrolepidópteros (Lepidoptera) del sureste de la península Ibérica.– Boletín de la Asociación Española de Entomología, 34(1-2): 123-143.
PEINADO, M., ALCARAZ, F. & MARTÍNEZ-PARRAS, J. M., 1992.– Vegetation of Southeastern Spain: 487 pp. J. Cramer, Berlin.
REDONDO, V. M., GASTÓN, F. J. & GIMENO, R., 2009.– Geometridae Ibericae: 361 pp. Apollo Books, Stenstrup.
REDONDO, V., GASTÓN, J. & VICENTE, J. C., 2010.– Las mariposas de la España Peninsular: 405 pp. Prames, Zaragoza.
ROBINEAU, R., 2007.– Guide des papillons nocturnes de France: 288 pp. Delachaux et Nietslé S. A., (eds.), Paris.
SKOU, P., 2007.– Menophra annegreteae sp. n., a new ennominae well established in southern Spain, with notes on the status of Sardocyrnia fortunaria (Vázquez, 1905) (Geometridae: Ennominae).– Nota lepidopterologica, 29: 137-144.
VALLHONRAT, F., 1980.– Pingasa lahayei Oberthür: geomètrid nord-africà recollit a Andalusia.– Treballs de la Societat Catalana de Lepidopterologia, 3: 67.
Familia Geometridae Leach, 1815
Subfamilia Ennominae Duponchel, 1845
Chiasmia aestimaria (Hübner, [1809]) Material estudiado: El Pueblo Indalo, 7-IV-2010, 1 ♂; 25-IV-2011, 1 ♂; 27-IV-2011, 1 ♂; Rambla de Morales, 22-IV-2014, 1 ♂. Citas bibliográficas: KRAUS (1999). Elemento asiático-mediterráneo. Bivoltina. Imagos: IV-V.
**Narraga nelvae (Rothschild, 1912) Material estudiado: Rambla de la Granatilla, 17-VI-2012, 1 ♂; El Algarrobico, 10-III-2013, 1 ♂. Elemento atlanto-mediterráneo. Bivoltina. Imagos: II-X.
Isturgia miniosaria (Duponchel, 1829) Material estudiado: El Saladero, 2-XI-2012, 1 ♂; 22-XI-2012, 1 ♂; El Algarrobico, 16-XII-2013, 1 ♂. Citas bibliográficas: KRAUS (1997, 1999). Elemento atlanto-mediterráneo. Univoltina. Imagos: IX-XII.
Itame vincularia (Hübner, [1813]) Material estudiado: El Saladero, 6-VIII-2012, 1 ♀. Citas bibliográficas: HACKER & WOLF (1982); KRAUS (1997, 1999). Elemento atlanto-mediterráneo. Bivoltina. Imagos: I-XI.
Acanthovalva inconspicuaria (Hübner, [1819]) Material estudiado: Rambla de la Granatilla, 8-VI-2009, 1 ♂, 1 ♀; El Saladero, 31-VII-2012, 1 ♂. Citas bibliográficas: KRAUS (1997, 1999). Elemento tropical. Polivoltina. Imagos: I-XII.
Isturgia catalaunaria (Guenée, 1858) Material estudiado: El Saladero, 31-VII-2012, 1 ♂; 8-VIII-2012, 1 ♂. Citas bibliográficas: HACKER & WOLF (1982); KRAUS (1999). Elemento tropical. Polivoltina. Imagos: I-XII.
Rhoptria asperaria (Hübner, [1817]) Material estudiado: El Saladero, 17-X-2012, 1 ♀; 4-II-2013, 1 ♂. Citas bibliográficas: KRAUS (1999). Elemento atlanto-mediterráneo. Bivoltina. Imagos: II-IV, VIII-XII.
**Petrophora narbonea (Linnaeus, 1767) Material estudiado: Rambla de la Granatilla, 21-III-2010, 1 ♂; El Saladero, 17-X-2012, 1 ♂; Playa del Charco, 8-XII-2013, 1 ♂; Cerro de la Cruz, 16-II-2014, 1 ♂. Elemento atlanto-mediterráneo. Bivoltina. Imagos: I-VI, IX-XII.
Petrophora convergata (Villers, 1789) Material estudiado: El Algarrobico, 7-X-2012, 1 ♂. Citas bibliográficas: MAGRO (2013). Elemento atlanto-mediterráneo. Univoltina. Imagos: X-XII.
Toulgoetia cauteriata (Staudinger, 1859) Material estudiado: El Algarrobico, 17-II-2013, 1 ♀; El Saladero, 16-III-2013, 1 ♂. Citas bibliográficas: KRAUS (1999). Elemento atlanto-mediterráneo. Univoltina. Imagos: II-III.
*Chemerina caliginearia (Rambur, 1833)Material estudiado: El Algarrobico, 22-XI-2012, 1 ♂; El Saladero, 22-XI-2012, 1 ♂ . Elemento atlanto-mediterráneo. Univoltina. Imagos: XI-III.
**Menophra abruptaria (Thunberg, 1792) Material estudiado: Rambla de la Granatilla, 21-III-2010, 1 ♂; El Saladero, 7-VIII-2012, 1 ♂; 17- X-2012, 1 ♂; 23-XII-2012, 3 ♂ ♂; 3-II-2014, 1 ♂; El Algarrobico, 22-XI-2012, 1 ♂; 9-XII-2012, 1 ♂; 23-XII-2012, 1 ♂; 2-IV-2013, 1 ♀. Elemento asiático-mediterráneo. Polivoltina. Imagos: I-VIII, X-XII.
Menophra japygiaria (Costa, 1849) Material estudiado: Rambla de la Granatilla, 20-III-2010, 1 ♂; 18-VIII-2010, 1 ♂; El Algarrobico, 3-VIII-2012, 1 ♂; El Saladero, 16-III-2013, 1 ♂. Citas bibliográficas: KRAUS (1997, 1999); MAGRO (2013). Elemento asiático-mediterráneo. Polivoltina. Imagos: I, III-IV, VI-VIII, X-XII.
Menophra annegreteae Skou, 2007 Material estudiado: El Saladero, 1-VII-2012, 1 ♂; 9-XII-2012, 1 ♂. Citas bibliográficas: KRAUS (1999) como Menophra undulosa (Albers & Warnecke, 1941); SKOU (2007); REDONDO et al. (2009). Elemento endémico. Polivoltina. Imagos: III, V-VII, X-XII.
Phaselia algiricaria Oberthür, 1913 Material estudiado: Rambla de la Granatilla, 6-IV-2010, 1 ♂; El Saladero, 3-VI-2012, 1 ♂; 15-VII- 2012, 1 ♂; 7-VIII-2012, 1 ♂; Río Alías, 19-V-2013, 1 ♂; Cerro de la Cruz, 17-IV-2014, 1 ♂. Citas bibliográficas: ORTÍZ et al. (2010). Elemento atlanto-mediterráneo. Bivoltina. Imagos: III-X.
*Calamodes occitanaria (Duponchel, 1829) Material estudiado: El Algarrobico, 2-XI-2012, 1 ♀; Playa del Charco, 8-XII-2013, 1 ♀. Elemento atlanto-mediterráneo. Univoltina. Imagos: X-XII.
Sardocyrnia fortunaria (Vázquez, 1905) Material estudiado: Rambla de la Granatilla, 20-III-2010, 1 ♂; 23-VIII-2010, 1 ♂; 26-IV-2011, 1 ♂; El Algarrobico, 27-I-2013, 1 ♂; Playa del Charco, 24-III-2014, 1 ♂. Citas bibliográficas: KRAUS (1997, 1999) como Sardocyrnia bastelicaria (Bellier, 1862). Elemento endémico. Polivoltina. Imagos: I-VI, VIII-X, XII.
*Selidosema taeniolaria (Hübner, [1813]) Material estudiado: El Algarrobico, 10-IX-2012, 1 ♂ ; 24-IX-2012, 1 ♀. Elemento atlanto-mediterráneo. Univoltina. Imagos: VIII-X.
*Tephronia codetaria (Oberthür, 1881) Material estudiado: Salinas de Cabo de Gata, 10-V-2013, 2 ♂ ♂; Rambla de Morales, 19-X-2013, 1 ♂; 2-V-2014, 1 ♂. Elemento atlanto-mediterráneo. Bivoltina. Imagos: V, X.
Lhommeia biskraria (Oberthür, 1885) Material estudiado: Playa del Charco, 19-X-2013, 1 ♂. Citas bibliográficas: MAGRO (2013). Elemento atlanto-mediterráneo. Polivoltina. Imagos: X.
**Gnophos perspersata Treitschke, 1827 Material estudiado: Rambla de la Granatilla, 20-V-2012, 1 ♂. Elemento atlanto-mediterráneo. Univoltina. Imagos: IV-VII.
Charissa mucidaria (Hübner, [1799]) Material estudiado: El Saladero, 22-XI-2012, 1 ♀; Cerro de la Cruz, 16-II-2014, 1 ♂. Citas bibliográficas: HACKER & WOLF (1982). Elemento atlanto-mediterráneo. Polivoltina. Imagos: I-V, IX-XII.
Charissa assoi (Redondo & Gastón, 1997) Citas bibliográficas: GARRE et al. (2013). Elemento endémico. Polivoltina. Imagos: VI, XI.
Aspitates ochrearia (Rossi, 1794) Material estudiado: Rambla de la Granatilla, 7-X-2012, 1 ♂; Río Alías, 17-II-2013, 1 ♂. Citas bibliográficas: HACKMAN (1968). Elemento euroasiático. Bivoltina. Imagos: II-IV, X.
Dyscia penulataria (Hübner, [1819]) Material estudiado: El Saladero, 31-VII-2012, 1 ♂. Citas bibliográficas: KRAUS (1997). Elemento atlanto-mediterráneo. Bivoltina. Imagos: I-XII.
Onychora agaritharia (Dardoin, 1842) Material estudiado: El Algarrobico, 24-IX-2012, 1 ♂; 17-X-2012, 1 ♂ ; Río Alías, 1-IX-2013, 1 ♂. Citas bibliográficas: KRAUS (1999). Elemento atlanto-mediterráneo. Univoltina. Imagos: IX-XI.
*Compsoptera opacaria (Hübner, [1819]) Material estudiado: El Saladero, 7-X-2012, 2 00; 1-XI-2013, 1 ♀. Elemento atlanto-mediterráneo. Univoltina. Imagos: X-XI.
Subfamilia Desmobathrinae Meyrick, 1886
**Myinodes interpunctaria (Herrich-Schäffer, 1839) Material estudiado: El Saladero, 4-II-2013, 1 ♂; 16-III-2013, 2 ♂ ♂; Río Alías, 17-II-2013, 1 ♂; Cañada del Madroñal, 16-II-2014, 3 ♂ ♂; 17-III-2014, 1 ♀. Elemento atlanto-mediterráneo. Univoltina. Imagos: II-III.
Subfamilia Geometrinae Stephens, 1829
Pingasa lahayei (Oberthür, 1887) Citas bibliográficas: VALLHONRAT (1980); KRAUS (1997, 1999). Elemento tropical. Bivoltina. Imagos: V, VIII.
Pseudoterpna coronillaria (Hübner, [1817]) Citas bibliográficas: ORTÍZ et al. (2010). Elemento asiático-mediterráneo. Bivoltina. Imagos: V-VI, X.
Thetidia plusiaria Boisduval, 1840 Material estudiado: El Algarrobico, 19-V-2013, 1 ♂; Rambla de Morales, 2-V-2014, 1 ♀. Citas bibliográficas: AGENJO (1964). Elemento atlanto-mediterráneo. Bivoltina. Imagos: IV-XI.
**Kuchleria insignata Hausmann, 1994 Material estudiado: Rambla de la Granatilla, 16-VIII-2010, 1 ♂; El Saladero, 17-VI-2012, 1 ♂; 2- IV-2013, 1 ♂; El Algarrobico, 20-XI-2013, 1 ♀. Elemento atlanto-mediterráneo. Polivoltina. Imagos: III-XI.
Phaiogramma etruscaria (Zeller, 1849) Material estudiado: Rambla de la Granatilla, 16-VIII-2010, 1 ♂; 19-VIII-2010, 1 ♀; El Algarrobico, 10-IX-2012, 1 ♂. Citas bibliográficas: HACKER & WOLF (1982) como Chlorissa pulmentaria (Guenée, 1857); MUÑOZ (1992). Elemento asiático-mediterráneo. Bivoltina. Imagos: IV-IX.
**Phaiogramma faustinata (Millière, 1868) Material estudiado: El Saladero, 3-VI-2012, 1 ♂; 17-VI-2012, 1 ♂; 17-II-2013, 1 ♂. Elemento asiático-mediterráneo. Polivoltina. Imagos: II-IX, XI.
Microloxia herbaria (Hübner, [1813]) Material estudiado: Rambla de la Granatilla, 20-VIII-2010, 1 ♀; El Saladero, 1-VII-2012, 1 ♀. Citas bibliográficas: AGENJO (1952). Elemento asiático-mediterráneo. Polivoltina. Imagos: IV-VIII, X.
Subfamilia Sterrhinae Meyrick, 1892
Idaea mediaria (Hübner, [1819]) Material estudiado: El Saladero, 23-VI-2013, 1 ♂. Citas bibliográficas: ORTÍZ et al. (2010). Elemento atlanto-mediterráneo. Univoltina. Imagos: IV-VII.
*Idaea ochrata (Scopoli, 1763) Material estudiado: El Algarrobico, 3-VI-2012, 1 ♂. Elemento asiático-mediterráneo. Univoltina. Imagos: IV-VI.
Idaea alicantaria (Reisser, 1963) Material estudiado: El Algarrobico, 3-VIII-2012, 1 ♀; Rambla de Morales, 10-V-2013, 1 ♀; 22-IV- 2014, 1 ♀; Río Alías, 19-V-2013, 1 ♂. Citas bibliográficas: KRAUS (1999); HAUSMANN (2004). Elemento endémico. Bivoltina. Imagos: IV-VI, VIII-X.
Idaea mustelata (Gumppenberg, 1892) Citas bibliográficas: ORTÍZ et al. (2010). Elemento atlanto-mediterráneo. Bivoltina. Imagos: VI-VII, X.
Idaea attenuaria (Rambur, 1933) Material estudiado: Rambla de la Granatilla, 7-IV-2010, 1 ♂, 1 ♀; 24-IX-2012, 1 ♂; El Algarrobico, 2-IV-2013, 1 ♂; 5-X-2013, 1 ♀; El Saladero, 8-V-2014, 1 ♂. KRAUS (1999) menciona la especie Idaea incalcarata (Chrétien, 1913) con la misma fecha de captura que, con anterioridad (Kraus, 1997), lo hace para I. attenuaria, aparentemente, corrigiendo esta última cita. Considerando que I. attenuaria está más vinculada a hábitats costeros (HAUSMANN, 2004) y es la única que se ha capturado en este estudio, parece más razonable mantener la referencia a este taxón. Citas bibliográficas: KRAUS (1997, 1999). Elemento atlanto-mediterráneo. Bivoltina. Imagos: III-V, IX-XI.
**Idaea alyssumata (Millière, 1871) Material estudiado: Rambla de la Granatilla, 23-VIII-2010, 1 ♀; El Saladero, 3-VIII-2012, 1 ♂; El Algarrobico, 16-IX-2013, 1 ♀. Elemento atlanto-mediterráneo. Bivoltina. Imagos: VIII-IX.
**Idaea urcinata (Agenjo, 1952) Material estudiado: Rambla de la Granatilla, 20-VIII-2010, 1 ♀; 20-V-2012, 1 ♂; 17-VI-2012, 1 ♀; Río Alías, 1-IX-2013, 1 ♀. Elemento endémico. Bivoltina. Imagos: V-VI, VIII-IX.
**Idaea rainerii Hausmann, 1994 Material estudiado: Rambla de la Granatilla, 18-VIII-2010, 1 ♂ ; 19-VIII-2010, 1 ♂; 21-VIII-2010, 1 ♀; 23-VIII-2010, 1 ♂ ; Río Alías, 19-V-2013, 1 ♂, 1 ♀. Elemento atlanto-mediterráneo. Bivoltina. Imagos: V-VI, VIII-IX.
Idaea calunetaria (Staudinger, 1859) Citas bibliográficas: HACKER & WOLF (1982). Elemento atlanto-mediterráneo. Bivoltina. Imagos: IX.
**Idaea belemiata (Millière, 1868) Material estudiado: El Algarrobico, 31-VII-2012, 1 ♀; 1-VIII-2012, 1 ♂, 2 ♀ ♀; 3-VIII-2012, 1 ♀. Elemento atlanto-mediterráneo. Univoltina. Imagos: VII-VIII.
Idaea elongaria (Rambur, 1833)Material estudiado: El Algarrobico, 15-VIII-2012, 1 ♂; El Saladero, 3-VIII-2012, 1 ♂. Citas bibliográficas: HACKER & WOLF (1982); KRAUS (1997, 1999). Elemento asiático-mediterráneo. Bivoltina. Imagos: V-IX.
**Idaea obsoletaria (Rambur, 1833) Material estudiado: El Saladero, 17-VI-2012, 1 ♀; 15-VII-2012, 1 ♀. Elemento asiático-mediterráneo. Univoltina. Imagos: VI-VIII.
Idaea blaesii Lenz & Hausmann, 1992 Citas bibliográficas: HAUSMANN (2004); REDONDO et al. (2009, 2010). Elemento atlanto-mediterráneo. Bivoltina.
Idaea lutulentaria (Staudinger, 1892) Citas bibliográficas: ORTÍZ et al. (2010). Elemento endémico. Univoltina. Imagos: VI.
Idaea longaria (Herrich-Schäffer, 1852) Material estudiado: Rambla de la Granatilla, 20-III-2010, 1 ♂; 21-III-2010, 1 ♂; 6-IV-2010, 1 ♂; El Algarrobico, 31-VII-2012, 1 ♂; 17-X-2012, 1 ♂; El Saladero, 2-XI-2012, 1 ♂. Citas bibliográficas: MUÑOZ (1992). Elemento asiático-mediterráneo. Polivoltina. Imagos: I-XII.
Idea minuscularia ( Ribe, 1912) Material estudiado: Rambla de la Granatilla, 20-III-2010, 1 ♂ ; 21-III-2010, 2 ♂ ♂ ; 6-IV-2010, 1 ♀; 17-VIII-2010, 1 ♂; El Algarrobico, 3-VII-2012, 1 ♂ ; El Saladero, 22-XI-2012, 1 ♂; 9-XII-2012, 1 ♀. Citas bibliográficas: MUÑOZ (1992) como Idaea herbuloti Agenjo, 1952; KRAUS (1999). Elemento atlanto-mediterráneo. Polivoltina. Imagos: I-VIII, X-XII.
Idaea carvalhoi Herbulot, 1979 Material estudiado: Rambla de la Granatilla, 9-VI-2009, 1 ♂; El Saladero, 1-VII-2012, 1 ♀; 3- VIII-2012, 2 ♀♀; 1-VI-2013, 1 ♀; El Algarrobico, 6-VIII-2012, 2 ♂ ♂; 10-IX-2012, 1 ♂. Citas bibliográficas: KRAUS (1999). Elemento atlanto-mediterráneo. Univoltina. Imagos: V-X.
Idaea subsericeata (Haworth, 1809) Material estudiado: Rambla de la Granatilla, 7-IV-2010, 2 ♂ ♂; El Algarrobico, 10-III-2013, 1 ♂; El Saladero, 2-IV-2013, 1 ♂. Citas bibliográficas: ORTÍZ et al. (2010). Elemento euroasiático. Bivoltina. Imagos: III-VI, VIII.
**Idaea fractilineata (Zeller, 1847) Material estudiado: Rambla de la Granatilla, 9-VI-2009, 1 ♂ ; 18-VIII-2010, 1 ♂; El Algarrobico, 24-IX-2012, 2 ♂ ♂; Salinas de Cabo de Gata, 10-V-2013, 1 0; Río Alías, 19-V-2013, 2 ♀ ♀; 1-VI-2013, 1 ♀; 8-V-2014, 1 ♂; Rambla de Morales, 2-V-2014, 1 ♀. Elemento atlanto-mediterráneo. Polivoltina. Imagos: V-IX.
Idaea cervantaria (Millière, 1869) Material estudiado: Rambla de la Granatilla, 20-III-2010, 1 ♂; 21-III-2010, 1♀; El Saladero, 1- VII-2012, 1 ♂; 9-XII-2012, 1 ♀; 8-I-2013, 1 ♀. Citas bibliográficas: HACKER & WOLF (1982). Elemento atlanto-mediterráneo. Polivoltina. Imagos: I-VI, IX-XII.
**Idaea deitanaria Reisser & Weisert, 1977 Material estudiado: Rambla de la Granatilla, 27-IV-2011, 1 ♂; 11-IV-2012, 2 ♂ ♂; El Saladero, 1- VII-2012, 2 ♂ ♂ ; 6-IV-2014, 1 ♂; El Algarrobico, 31-VII-2012, 1 ♀; 2-IV-2013, 1 ♂. Elemento endémico. Bivoltina. Imagos: IV-XI.
*Idaea infirmaria (Rambur, 1833) Material estudiado: El Saladero, 17-VI-2012, 1 ♀; 15-VII-2012, 1 ♂. Elemento atlanto-mediterráneo. Univoltina. Imagos: VI-IX.
Idaea ostrinaria (Hübner, [1813]) Material estudiado: Río Alías, 16-IV-2013, 1 ♂. Citas bibliográficas: ORTÍZ et al. (2010). Elemento asiático-mediterráneo. Univoltina. Imagos: IV-VI.
Idaea eugeniata (Dardoin & Millière, 1870) Material estudiado: Rambla de la Granatilla, 20-III-2010, 1 ♂; 21-VIII-2010, 1 ♀; El Saladero, 17- X-2012, 1 ♂; 16-III-2013, 1 ♂; 2-IV-2013, 1 ♀. Citas bibliográficas: HACKER & WOLF (1982). Elemento atlanto-mediterráneo. Polivoltina. Imagos: III-X.
*Idaea degeneraria (Hübner, [1799]) Material estudiado: El Algarrobico, 6-IV-2014, 1 ♀. Elemento asiático-mediterráneo. Polivoltina. Imagos: IV.
Brachyglossina hispanaria (Püngeler, 1913) Material estudiado: Rambla de la Granatilla, 18-VIII-2010, 1 ♂; El Saladero, 17-VI-2012, 1 ♀; Río Alías, 2-V-2013, 1 ♂. Citas bibliográficas: ORTÍZ et al. (2010). Elemento endémico. Univoltina. Imagos: IV-IX.
Oar reaumuraria (Millière, 1864) Material estudiado: Rambla de Morales, 10-V-2013, 2 ♂ ♂; 2-V-2014, 1 ♂, 1 ♀; El Algarrobico, 19-V-2013, 1 ♂; Playa del Charco, 19-X-2013, 1 ♂. Citas bibliográficas: Como Oar pratana (Fabricius, 1794) en HACKER & WOLF (1982), MUÑOZ (1992) y KRAUS (1997, 1999). Elemento atlanto-mediterráneo. Polivoltina. Imagos: III-V, VIII-XI.
Cinglis andalusiaria Wagner, 1935 Material estudiado: Rambla de la Granatilla, 20-VIII-2010, 1 ♂; 2-VIII-2012, 1 ♀. Citas bibliográficas: MUÑOZ (1992) como Cinglis humifusaria (Eversmann, 1837). Elemento atlanto-mediterráneo. Bivoltina. Imagos: IV-IX.
**Scopula submutata (Treitschke, 1828) Material estudiado: Rambla de Macenas, 26-IV-2011, 1 ♀. Elemento asiático-mediterráneo. Bivoltina. Imagos: IV-VIII, X.
**Scopula decolor (Staudinger, 1898) Material estudiado: El Algarrobico, 27-I-2013, 1 ♂ ; 3-II-2014, 1 ♂; Rambla de Morales, 24-III- 2014, 1 ♂, 1 ♀; 22-IV-2014, 1 ♀. Elemento asiático-mediterráneo. Polivoltina. Imagos: I-IV.
**Scopula marginepunctata (Goeze, 1781) Material estudiado: El Saladero, 1-VII-2012, 1 ♀; El Algarrobico, 8-VIII-2012, 1 ♂. Elemento euroasiático. Polivoltina. Imagos: IV-VIII, XII.
Scopula imitaria (Hübner, [1799]) Material estudiado: Rambla de la Granatilla, 26-IV-2011, 1 ♂; El Saladero, 9-XII-2012, 1 ♂; El Algarrobico, 4-II-2013, 1 ♂; 10-III-2013, 1 ♂. Citas bibliográficas: KRAUS (1999). Elemento asiático-mediterráneo. Polivoltina. Imagos: II-VIII, XII.
Scopula minorata (Boisduval, 1833) Material estudiado: El Pueblo Indalo, 30-IV-2009, 1 ♀; 7-VI-2009, 1 ♂. Citas bibliográficas: HACKER & WOLF (1982). Elemento cosmopolita. Polivoltina. Imagos: IV-VI, VIII-IX.
Glossotrophia asellaria (Herrich-Schäffer, 1849) Material estudiado: Rambla de la Granatilla, 21-III-2010, 1 ♂, 2 ♀♀; El Algarrobico, 2-XI-2012, 1 ♀. Citas bibliográficas: KRAUS (1997, 1999). Elemento atlanto-mediterráneo. Polivoltina. Imagos: I-XII.
**Rhodostrophia pudorata (Fabricius, 1794) Material estudiado: El Algarrobico, 20-V-2012, 1 ♂; Río Alías, 2-V-2013, 1 ♂. Elemento atlanto-mediterráneo. Univoltina. Imagos: IV-VI.
**Rhodostrophia calabra (Petagna, 1786) Material estudiado: Rambla de la Granatilla, 26-IV-2011, 1 ♂; El Algarrobico, 16-IV-2013, 1 ♂. Elemento asiático-mediterráneo. Univoltina. Imagos: IV-VI.
Cyclophora puppillaria (Hübner, [1799]) Material estudiado: Rambla de la Granatilla, 16-VIII-2010, 1 ♂; El Saladero, 10-III-2013, 1 ♂; El Algarrobico, 5-X-2013, 1 ♀. Citas bibliográficas: ORTÍZ et al. (2010). Elemento asiático-mediterráneo. Polivoltina. Imagos: III-IV, VI-VIII, X-XI.
**Rhodometra sacraria (Linnaeus, 1767) Material estudiado: Rambla de la Granatilla, 7-IV-2010, 1 ♀; El Saladero, 24-IX-2012, 1 ♂. Elemento cosmopolita. Polivoltina. Imagos: III-XI.
Casilda consecraria (Staudinger, 1871) Material estudiado: El Saladero, 2-XI-2012, 1 ♂; Salinas de Cabo de Gata, 10-V-2013, 1 ♀; Rambla de Morales, 19-X-2013, 1 ♂; 22-IV-2014, 1 ♀. Citas bibliográficas: KRAUS (1999). Elemento euroasiático. Polivoltina. Imagos: IV-V, X-XI.
Subfamilia Larentiinae Duponchel, 1845
**Scotopteryx peribolata (Hübner, [1817]) Material estudiado: El Algarrobico, 7-X-2012, 1 ♂; 5-X-2013, 1 ♂. Elemento atlanto-mediterráneo. Univoltina. Imagos: X.
Orthonama obstipata (Fabricius, 1794) Material estudiado: El Algarrobico, 8-VII-2013, 1 ♂. Citas bibliográficas: ORTÍZ et al. (2010). Elemento cosmopolita. Polivoltina. Imagos: VI-VII.
*Xanthorhoe fluctuata (Linnaeus, 1758) Material estudiado: Rambla de la Granatilla, 6-IV-2010, 1 ♂; El Saladero, 17-X-2012, 1 ♂; El Algarrobico, 22-XI-2012, 1 ♂; Cerro de la Cruz, 16-II-2014, 1 ♂. Elemento paleártico. Polivoltina. Imagos: I-V, X-XII.
*Catarhoe basochesiata (Duponchel, 1861) Material estudiado: Rambla de la Granatilla, 7-IV-2010, 1 ♀; 9-XII-2012, 1 ♂; El Saladero, 2-XI- 2012, 2 ♂ ♂; El Algarrobico, 2-XI-2012, 1 ♀; 17-II-2013, 1 ♀. Elemento atlanto-mediterráneo. Bivoltina. Imagos: I-IV, X-XII.
**Epirrhoe sandosaria (Herrich-Schäffer, 1852) Material estudiado: Rambla de la Granatilla, 21-III-2010, 1 ♀; 7-IV-2010, 1 ♂, 1 ♀; 10-IV-2012, 1 ♂; El Saladero, 17-X-2012, 1 ♀; 9-XII-2012, 1 ♀. Elemento atlanto-mediterráneo. Bivoltina. Imagos: I-V, X-XII.
Larentia malvata (Rambur, 1833) Material estudiado: El Saladero, 17-X-2012, 1 ♂ ; 2-XI-2012, 1 ♂ , 1 ♀; El Algarrobico, 2-XI-2012, 1 ♂, 1 0; 22-XI-2012, 1 ♂; 9-XII-2012, 1 ♂; 20-XI-2013, 1 ♂, 1 ♀; 16-XII-2013, 1 ♀. Citas bibliográficas: HAUSMANN & VIIDALEPP (2012). Elemento atlanto-mediterráneo. Univoltina. Imagos: X-I.
Antilurga alhambrata (Staudinger, 1859) Aunque se trata de una especie de vuelo típicamente otoñal (HAUSMANN & VIIDALEPP, 2012), solo ha sido citada en el mes de abril. Citas bibliográficas: KRAUS (1997, 1999). Elemento atlanto-mediterráneo. Univoltina. Imagos: IV.
*Nebula ibericata (Staudinger, 1871) Material estudiado: Rambla de la Granatilla, 6-IV-2010, 1 ♂; El Algarrobico, 17-X-2012, 1 ♂. Elemento atlanto-mediterráneo. Bivoltina. Imagos: I-IV, X-XII.
*Colostygia multistrigaria (Haworth, 1809) Material estudiado: El Saladero, 23-XII-2012, 1 ♂; El Algarrobico, 23-XII-2012, 1 ♂. Elemento asiático-mediterráneo. Univoltina. Imagos: XII-I.
**Almeria kalischata (Staudinger, 1870) Material estudiado: Rambla de la Granatilla, 9-VI-2009, 1 ♀; 20-III-2010, 1 ♀; El Algarrobico, 23- XII-2012, 1 ♂. Elemento atlanto-mediterráneo. Bivoltina. Imagos: I-IX, XI-XII.
*Hospitalia flavolineata (Staudinger, 1883) Material estudiado: El Saladero, 7-X-2012, 1 ex. Elemento atlanto-mediterráneo. Univoltina. Imagos: X.
Horisme scorteata (Staudinger, 1901) Material estudiado: Rambla de la Granatilla, 20-III-2010, 1 ♀; 21-III-2010, 1 ♀; 6-IV-2010, 1 ♂; 7- IV-2010, 1 ♂, 1 ♀. Citas bibliográficas: REDONDO et al. (2009). Elemento atlanto-mediterráneo. Polivoltina. Imagos: III-IV, X.
*Epirrita dilutata ([Denis & Schiffermüller], 1775) Material estudiado: Rambla de la Granatilla, 9-XII-2012, 1 ♂. Elemento asiático-mediterráneo. Univoltina. Imagos: XII.
Gymnoscelis rufifasciata (Haworth, 1809) Material estudiado: Rambla de la Granatilla, 9-XII-2012, 1 ♀; El Saladero, 4-II-2013, 1 ♀; 17-II- 2013, 1 ♂. Citas bibliográficas: AGENJO (1964); KRAUS (1997). Elemento paleártico. Polivoltina. Imagos: I-VIII, X-XII.
Eupithecia ultimaria Boisduval, 1840 Material estudiado: Rambla de la Granatilla, 6-IV-2010, 1 ♂; 19-VIII-2010, 1 ♂, 2 00; 21-VIII- 2010, 1 ♀; 10-IV-2012, 1 ♀; El Algarrobico, 8-VIII-2012, 1 ♀; Rambla de Morales, 22-IV-2014, 1 ♂, 1 ♀. Citas bibliográficas: MUÑOZ (1992); KRAUS (1997, 1999). Elemento asiático-mediterráneo. Bivoltina. Imagos: III-V, VIII-X.
Eupithecia minusculata Alphéraky, 1882 Material estudiado: Rambla de Morales, 2-V-2014, 1 ♂, 1 ♀. Citas bibliográficas: KRAUS (1999) como Eupithecia herrenschmidti von Mentzen & Moberg, 1992. Elemento asiático-mediterráneo. Polivoltina. Imagos: III, V.
*Eupithecia venosata (Fabricius, 1787) Material estudiado: Rambla de la Granatilla, 7-IV-2010, 1 ♀; Río Alías, 17-II-2013, 1 ♀; El Algarrobico, 17-II-2013, 1 ♂; 3-III-2014, 1 ♂. Elemento euroasiático. Univoltina. Imagos: II-IV.
*Eupithecia cocciferata Millière, 1864 Material estudiado: Rambla de la Granatilla, 20-III-2010, 1 ♂; El Algarrobico, 16-III-2013, 1 ♀; El Saladero, 16-III-2013, 1 ♀. Elemento atlanto-mediterráneo. Univoltina. Imagos: III-IV.
*Eupithecia dodoneata Guenée, 1858 Material estudiado: El Saladero, 4-II-2013, 1 ♀ ; 17-II-2013, 1 ♀; 10-III-2013, 1 ♀; 16-III-2013, 2 ♂ ♂; 2-IV-2013, 1 ♂, 1 ♀; 3-II-2014, 1 ♂; El Algarrobico, 4-II-2013, 1 ♂, 1 ♀; 16-III-2013, 1 ♀. Elemento euroasiático. Univoltina. Imagos: II-IV.
**Eupithecia massiliata Dardoin & Millière, 1865 Material estudiado: Rambla de la Granatilla, 7-IV-2010, 1 ♀. Elemento atlanto-mediterráneo. Univoltina. Imagos: IV.
Eupithecia oxycedrata (Rambur, 1833) Citas bibliográficas: HACKER & WOLF (1982). Elemento asiático-mediterráneo. Bivoltina. Imagos: IX.
**Eupithecia orana Dietze, 1804 Material estudiado: Rambla de la Granatilla, 20-III-2010, 1 ♂, 2 ♀ ♀; 9-XII-2012, 1 ♀; El Saladero, 17-X-2012, 1 ♂, 1 ♂; 22-XI-2012, 1 ♂; 17-II-2013, 1 ♂; El Algarrobico, 17-X-2012, 1 ♂, 1 ♀; 27-I- 2013, 1 ♂. Citas bibliográficas: EXPÓSITO-HERMOSA (1985). Elemento atlanto-mediterráneo. Bivoltina. Imagos: I-III, X-XII.
Eupithecia unedonata Mabille, 1868 Material estudiado: Rambla de la Granatilla, 21-III-2010, 1 ♀; 10-IV-2012, 1 ♂; El Algarrobico, 2- XI-2012, 1 ♂; El Saladero, 17-II-2013, 1 ♀; Playa del Charco, 24-III-2014, 1 ♂. Citas bibliográficas: KRAUS (1997, 1999). Elemento asiático-mediterráneo. Bivoltina. Imagos: II-IV, X-XII.
**Eupithecia unitaria Herrich-Schäffer, 1852 Material estudiado: Rambla de la Granatilla, 7-IV-2010, 2 ♂ ♂, 1 ♀; El Saladero, 2-IV-2013, 1 ♀. Elemento atlanto-mediterráneo. Bivoltina. Imagos: II-IV.
Eupithecia distinctaria Herrich-Schäffer, 1848 Material estudiado: Rambla de la Granatilla, 7-IV-2010, 3 ♂ ♂, 1 ♀; El Algarrobico, 20-V-2012, 1 ♂; 9-XII-2012, 2 00; 2-IV-2013, 1 ♂; El Saladero, 17-X-2012, 1 ♂; 19-V-2013, 1 ♂; 16-II-2013, 1 ♂; Río Alías, 1-VI-2013, 1 ♂ ; Cerro de la Cruz, 16-II-2014, 1 ♂. Citas bibliográficas: KRAUS (1999). Elemento euroasiático. Bivoltina. Imagos: I-VI, IX-XII.
*Eupithecia centaureata ([Denis & Schiffermüller], 1775) Material estudiado: El Saladero, 2-IV-2013, 1 ♂; Río Alías, 5-X-2013, 1 ♂; Cerro de la Cruz, 16- II-2014, 1 ♂. Elemento paleártico. Polivoltina. Imagos: II-VI, X-XI.
*Eupithecia variostrigata Alphéraky, 1876 Material estudiado: El Saladero, 20-XI-2013, 1 ♂. Elemento asiático-mediterráneo. Univoltina. Imagos: XI.
**Eupithecia gratiosata Herrich-Schäffer, 1861 Material estudiado: Rambla de Alfaix, 11-IV-2012, 1 ♂. Elemento euroasiático. Univoltina. Imagos: IV.
**Eupithecia semigraphata Bruand, 1850 Material estudiado: El Saladero, 17-X-2012, 1♀; El Algarrobico, 1-XII-2013, 2 ♀♀. Elemento asiático-mediterráneo. Bivoltina. Imagos: X-XI.
*Chesias isabella Schawerda, 1915 Material estudiado: Rambla de la Granatilla, 7-IV-2010, 1 ♀; 9-XII-2012, 1 ♂; El Saladero, 22-XI- 2012, 4 ♂ ♂; 9-XII-2012, 1 ♀; 23-XII-2012, 2 ♂♂; 4-II-2013, 2 ♂♂, 1 ♀; Río Alías, 3-II-2014, 1 ♀. Elemento atlanto-mediterráneo. Univoltina. Imagos: XI-IV.
Notas de autor
aortiz@um.es