
Eidophasia infuscata Staudinger, [1871] 1870, status nova with the first description of male and female genitalia (Lepidoptera: Plutellidae)
Eidophasia infuscata Staudinger, [1871] 1870, nuevo estatus con la primera descripción de la genitalia del macho y de la hembra (Lepidoptera: Plutellidae)
Eidophasia infuscata Staudinger, [1871] 1870, status nova with the first description of male and female genitalia (Lepidoptera: Plutellidae)
SHILAP Revista de lepidopterología, vol. 44, núm. 174, 2016
Sociedad Hispano-Luso-Americana de Lepidopterología
Recepción: 26 Noviembre 2014
Aprobación: 28 Diciembre 2014
Abstract: Eidophasia infuscata Staudinger from Greece, previously regarded as an infrasubspecies or a subspecies of Eidophasia messingiella, is proposed as a full species based on the differences in the coloration of the head, the forewing patterns, and the male and female genitalia. Eidophasia infuscata is reported from Turkey for the first time. The first descriptions of the genitalia of both sexes are provided for E. infuscata. A diagnostic summary is provided with illustrations for three similar congeners, Eidophasia messingiella, E. infuscata, and E. albifasciata.
Keywords: Lepidoptera, Plutellidae, Eidophasia infuscata, redescription, taxonomy, Greece.
Resumen: Eidophasia infuscata Staudinger de Grecia, considerada hasta ahora como una infrasubespecie o una subespecie de Eidophasia messingiella, se propone como una nueva especie, basándose en las diferencias en la coloración de la cabez, el diseño de las alas anteriores y las genitalias del macho y de la hembra. Eidophasia infuscata se cita de Turquía por primera vez y se aportan las primeras descripciones de las genitalias de ambos sexos. Se aporta también un diagnóstico de tres especies similares, Eidophasia messingiella, E. infuscata y E. albifasciata.
Palabras clave: Lepidoptera, Plutellidae, Eidophasia infuscata, redescripcion, taxonomía, Grecia.
Introduction
The plutellid genus Eidophasia currently comprises 11 species, most of which occur in the Holarctic Region (ROBINSON & SATTLER, 2001). Many species of Eidophasia are locally restricted, but one congener, E. messingiella (Fisher von Röslerstamm) is widespread across the Palearctic Region from Europe to Korea (SONH, 2012). Eidophasia messingiella include a few unique variations in the forewing patterns, some of which were considered as infrasubspecies (STAUDINGER, 1871; SCHILLE, 1907; CARADJA, 1920; DOETS, 1950). STAUDINGER (1871) noticed that E. messingiella from Greece differ from the conspecific populations from other European countries in the forewing median line. Regarding this difference, he described the Greek populations as a variety of E. messingiella, namely var. infuscata. FRIESE (1966) noted that the larvae of subspecies infuscata feed on Aubretia, differing from those of the nominal messingiella which feed on Cardamine. Despite these differences, Eidophasia messingiella var. infuscata has never been regarded as a separate species from E. messingiella (ROBINSON & SATTLER, 2001).
The aims of this article are to clarify the taxonomic status of Eidophasia messingiella var. infuscata and to re-describe it with male and female genitalia. Diagnostic differences between three confusing species, E. messingiella, E. infuscata, and E. albifaciata Issiki, are provided.
Materials and methods
Specimens were obtained from the following institutions.
MNHU: Museum für Naturkunde, Berlin, Germany;
USNM: United States National Museum of Natural History, Wasington, DC, USA;
ZMC: Zoologisk Museum, Copenhagen, Denmark;
ZSM: Zoologische Staatssammlung München, Munich, Germany.
Genitalia slides were prepared following CLARKE (1941), except that Chlorazol black was used for staining and Euparal resin for permanent mounting. Terms for genitalia follow KLOTS (1970). Verbatim label data are given for the lectotype of Eidophasia messingiella var. infuscata. The mark “ | ” in the label data indicates line breaks; additional information given by the present authors is provided in brackets. The ‘GSN’ stands for the genitalia slide number.
Systematic accounts
Eidophasia infuscata Staudinger, [1871] 1870, stat. nov. (Figs. 3-7, 10, 15)Eidophasia messingiella var. infuscataStaudinger, [1871] 1870: 238. Type locality: Greece, Mount Parnassus [central Greece near Delphi, north of the Gulf of Corinth, elevation 2457 m, prominence 1590 m, coordinates 38º 32’ 09”N, 22º 37’ 27”E].
Eidophasia messingiella ssp. infuscata; Friese, 1966: 450.
Eidophasia messingiella; Meyrick, 1914: 48;Robinson & Sattler, 2001: 25.
Types: Lectotype (designated here): male, “Origin” [pink label], “Graecia | Kr.” [yellow label; hand-written], “Parnass | 12/5 66.” [hand-written], “ex coll. 5/5 | STAUDINGER”, “Zool. Mus. | Berlin” [pale yellow label], “U. Roesler | 1 | GU: 5083”, “infuscala [sic] Sgr.” [hand-written with blue ink], deposited in MNHU. Paralectotypes (1 1, 3 00, 1 ex.): GREECE, Delphi, Mount Parnassus, 1 0, 12 May 1866, GSN: A 215; 1 1, 1 0, 13 May 1866; Macedonia, 1 ex., no date (Staudinger), all in MNHU.
Materials examined: GREECE, Peloponnese, Mts. Aroania (= Chelmos), 1400 m, 2 11, 10-VI- 1958 (J. Klimesch), reared from Aubrieta deltoidea (= A. intermedia), GSN: SJC-715, coll. ZSM; Kalavryta, Zachlorou, 1 0, 13-30-VI-1958 (J. Klimesch), coll. ZSM; Central Macedonia, Litochoro, 300-400 m, 1 0, 14-22-VI-1957 (J. Klimesch), GSN: SJC-716, coll. ZSM; Macedonia, Thessaly, Mount Olympus, 700-2100 m, 1 1, 21-26-V-1990, coll. ZMC. TURKEY, Kazi Bell, Denizli, Mount Honaz, 1250 m, 1 1, 1 0, 9-VI-1964 (J. Klimesch), coll. ZSM.
Redescription: Head - Vertex and frons dark brownish gray or dark brown. Antenna 2/3 as long as forewing; scape dark brownish gray, with awning forward; flagellomeres dark brown, with denser scales on first five flagellomeres in males and all but last 9-10 flagellomeres in females; most of the remaining flagellomeres with white annulation distally. Labial palpus slightly upcurved to middle of frons; 1st segment dark brown dorsally, white ventrally, 1/2 as long as 2nd; 2nd segment as long as 3rd, dark brown, tinged with white ventrobasally, with dark brown, elongate-scale tuft ventrally; 3rd segment dark brown on outer surface, pale brownish gray on inner surface. Thorax - Patagium, tegula, and mesonotum dark brown. Fore- and midleg with coxa, femur and tibia dark brown; tarsomere dark brown, with a pale yellow ring distally. Hindleg with coxa and femur dark brownish gray; tibia pale brownish gray; tarsomere dark brown with a pale yellow ring distally. Forewing length 5.1-6.5 mm (n = 4), dark brown, densely irrotated with brownish gray scales after antemedian line; termen more oblique in female; antemedian line nearly straight, pale yellow, of variable width, depending on individuals, inner margin sharp, outer margin diffused to pale orange; fringe dark brown, tinged with brownish gray basally. Hindwing and fringe brownish gray. Abdomen - Tergites and sternites dark brownish gray.
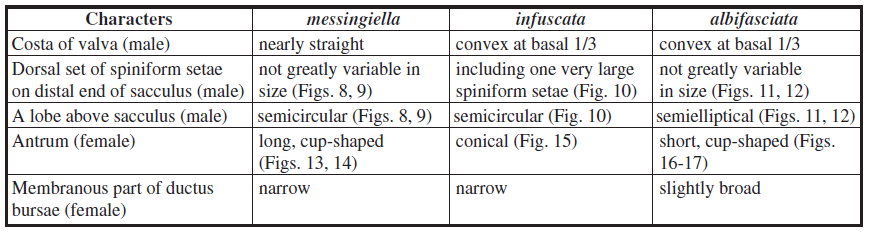
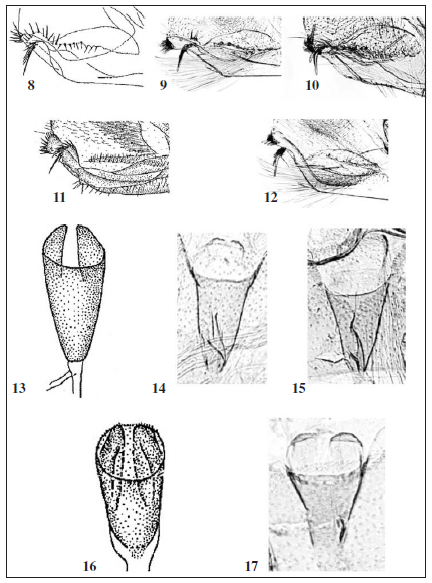
Male genitalia (Figs. 5-6): Tegumen semi-elliptical, with a pair of processes (‘teguminal process’) on apical 1/4; teguminal process right-subtriangular, setose, with acute projection apically. Tuba analis 1.4x longer than tegumen, with weak subscapium. Valva obovate, setose, with dense setal zone around apex; costa slightly convex at basal 1/3; a semicircular lobe above sacculus with spiniform setae in variable size along ventral margin; sacculus curved medially, broadened in basal half, with two sets of spiniform setae distally, dorsal set comprising one large and several short spiniform setae, ventral set comprising four curved spiniform setae in variable size and several short ones; a row of long setae along the basal 2/3 of sacculus. Vinculum triangular; saccus short, narrowly round apically. Phallus slightly curved at middle, slightly broadened in distal 1/6, bulbous basally. Female abdomen VIII with a zone of swirling wrinkles anterolaterally; caudal margin of sternite emarginated medially.
Female genitalia (Fig. 7): Papilla analis broad, subtriangular, setose. Abdomen IX with long setae along posterolateral margin and a pair of setose lobes ventromedially. Apophysis posterioris 3.2x longer than apophysis anterioris. A pair of semicircular, shallow depressions above ostium bursae. Ductus bursae with conical antrum in posterior half, slender in anterior half; inception of ductus seminalis near antrum; bulla seminalis ovobate, as long as corpus bursae. Corpus bursae elliptical, as long as antrum.
Distribution: Greece and Turkey.
Host plant: Brassicaceae, Aubrieta deltoidea (L.) DC.
Discussion
STAUDINGER (1870) noticed a few differences between Eidophasia messingiella and E. infuscata, including the coloration of the labial palpus, the ground color of the forewing, and the forewing patterns. My examination of additional specimens of E. infuscata revealed that the difference from E. messingiella in the forewing patterns is stable. The forewing median line of E. messingiella can be variable in width and continuity, but always has a sharp outer margin. In E. infuscata, the character is not present and instead, the outer margin of the forewing median line becomes gradually worn off. Further, the dark brown head of E. messingiella is tinged with pale brown along the lateral edges of the vertex. Differing from this, the vertex of the head of E. infuscata is entirely dark brown. More importantly, there are a few differences in the genital structures between E. messingiella and E. infuscata (Table 1). Regarding all these differences, both are better regarded as two distinctive species. Therefore, the records of E. messingiella from Greece (e.g. KARSHOL & RAZOWSKI, 1996) need to be reconsidered. Another congener, E. albifaciata, occurs in Japan and Korea (SOHN, 2012). The characters in the forewing patterns alone make it feasible to distinguish three species, E. messingiella, E. infuscata, and E. albifasciata. The genital features further support their distinctiveness as summarized in Table 1.

Agradecimientos
We would like to thank Donald Davis (United States National Museum of Natural History, Washington, DC), Ulf Buchsbaum, Axel Hausmann (both, Zoologische Staatssammlung München, Munich), Ole Karsholt (Zoologisk Museum, Copenhagen, Denmark), and Wolfram Mey (Museum für Naturkunde, Humboldt-Universität, Berlin) for allowing me to examine the institutional collections under their responsibility. This study was supported by the Ministry of Sciences and Higher Education (grant no. NN303 568538).
BIBLIOGRAPHY
CARADJA, A., 1920. – Beitrag zur Kenntnis der geographischen Verbreitung der Mikrolepidopteren des palaearktischen Faunengebietes nebst Beschreibung neuer Formen. 3.– Deutsche Entomologische Zeitschrift, Iris, 34: 75-179.
CLARKE, J. F. G.,1941 .– The preparation of slides of the genitalia of Lepidoptera.– Bulletin of the Brooklyn Entomological Society, 36: 149-161.
DOET, C., 1950.– Biology and variation of Eidophasia messingiella F. R. (Lep., Plutellidae).– Entomologische Berichten, Amsterdam, 13: 85-86.
FRIESE, G., 1966.– Ergebnisse der Albanien-Expedition 1961 des Deutschen Entomologischen Institutes. 52. Beitrag, Lepidoptera: Plutellidae.– Beiträge zur Entomologie, 16: 447-459.
KLOTS, A. B., 1970.– Lepidoptera.– In S. L. TUXEN (ed.).– Taxonomist’s Glossary of Genitalia in Insects: 115- 130. Munksgaard, Copenhagen.
MEYRICK, E., 1914.– Hyponomeutidae, Plutellidae, Amphitheridae.– In H. WAGNER (ed.).– Lepidopterorum Catalogus, Pars 19: 64 pp. W. Junk, Berlin.
KARSHOLT, O. & RAZOWSKI, J., 1996.– The Lepidoptera of Europe: 380 pp. Apollo Books, Stenstrup.
ROBINSON, G. & SATTLER, K., 2001.– Plutella in the Hawaiian Islands: relatives and host-races of the Diamondback Moth (Lepidoptera: Plutellidae).– Bishop Museum Occasional Papers, 67: 1-27.
SCHILLE, F., 1905.– Fauna lepidopterologiczna doliny Popradu i jego doplywów. VII- Sprawzdanie Komisyi Fizyograficznej, 38: 3-6.
SOHN, J.-C., 2012.– New records of Eidophasia messingiella (Fisher von Röslerstamm) from Korea and China (Yponomeutoidea, Plutellidae) with comments about forewing pattern variation in two East Asian Eidophasia.– Japan Heterocerists’ Journal, 265: 381-383.
STAUDINGER, O., [1871] 1870.– Beitrag zur Lepidopterenfauna Griechenlands.– Horae Societatis Entomologicae Rossicae, .: 3-304.
Notas de autor